

Directed differentiation and functional maturation of cortical interneurons from human embryonic stem cells
- PMID: 23642365
- PMCID: PMC3681523
- DOI: 10.1016/j.stem.2013.04.008
Directed differentiation and functional maturation of cortical interneurons from human embryonic stem cells
Abstract
Human pluripotent stem cells are a powerful tool for modeling brain development and disease. The human cortex is composed of two major neuronal populations: projection neurons and local interneurons. Cortical interneurons comprise a diverse class of cell types expressing the neurotransmitter GABA. Dysfunction of cortical interneurons has been implicated in neuropsychiatric diseases, including schizophrenia, autism, and epilepsy. Here, we demonstrate the highly efficient derivation of human cortical interneurons in an NKX2.1::GFP human embryonic stem cell reporter line. Manipulating the timing of SHH activation yields three distinct GFP+ populations with specific transcriptional profiles, neurotransmitter phenotypes, and migratory behaviors. Further differentiation in a murine cortical environment yields parvalbumin- and somatostatin-expressing neurons that exhibit synaptic inputs and electrophysiological properties of cortical interneurons. Our study defines the signals sufficient for modeling human ventral forebrain development in vitro and lays the foundation for studying cortical interneuron involvement in human disease pathology.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
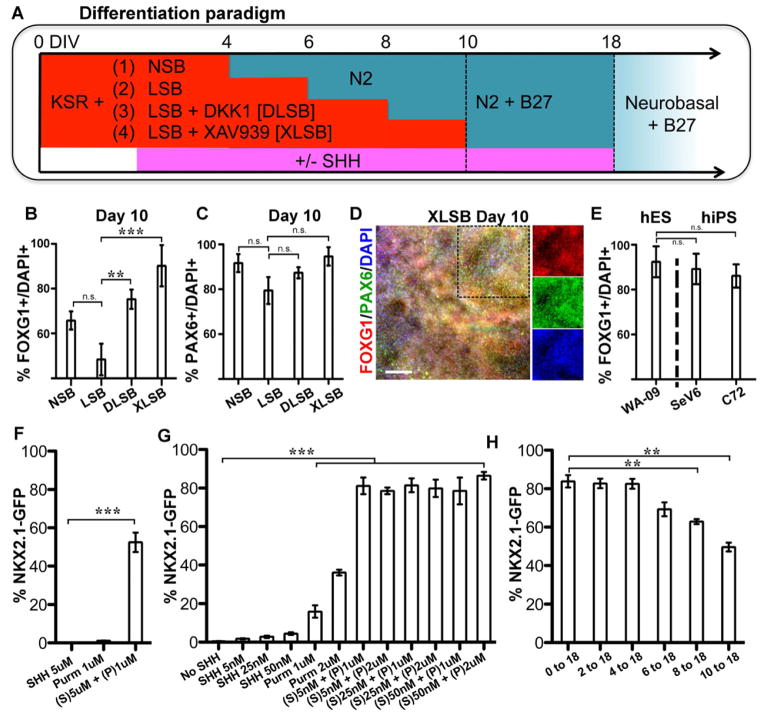
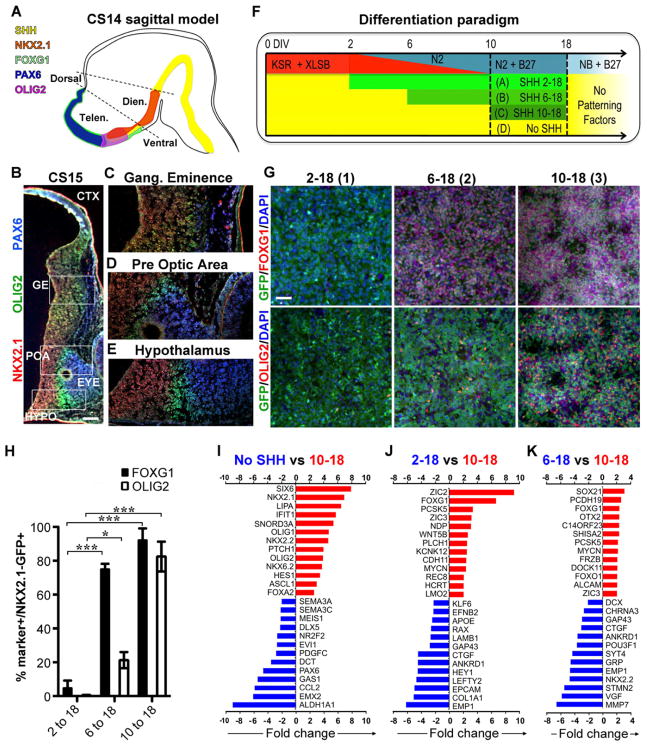
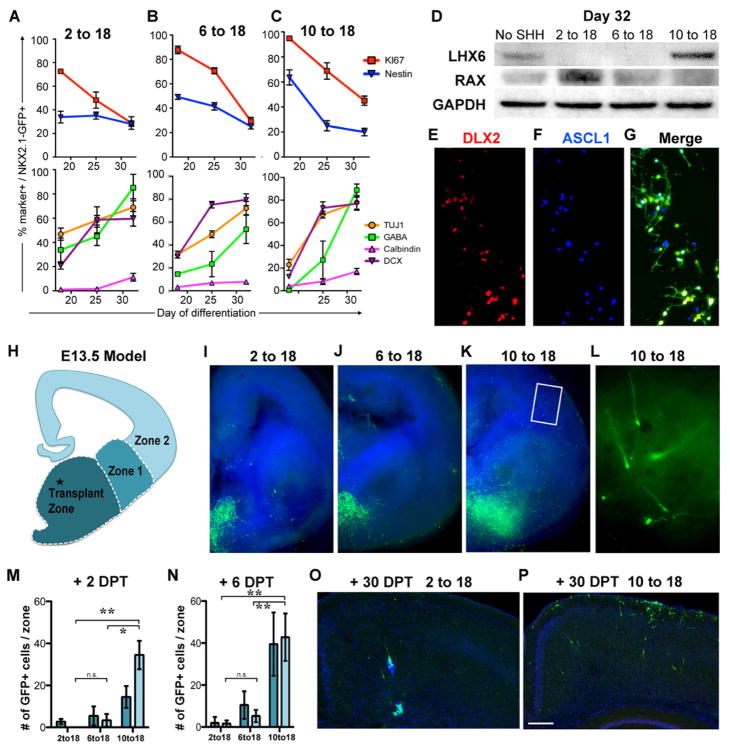
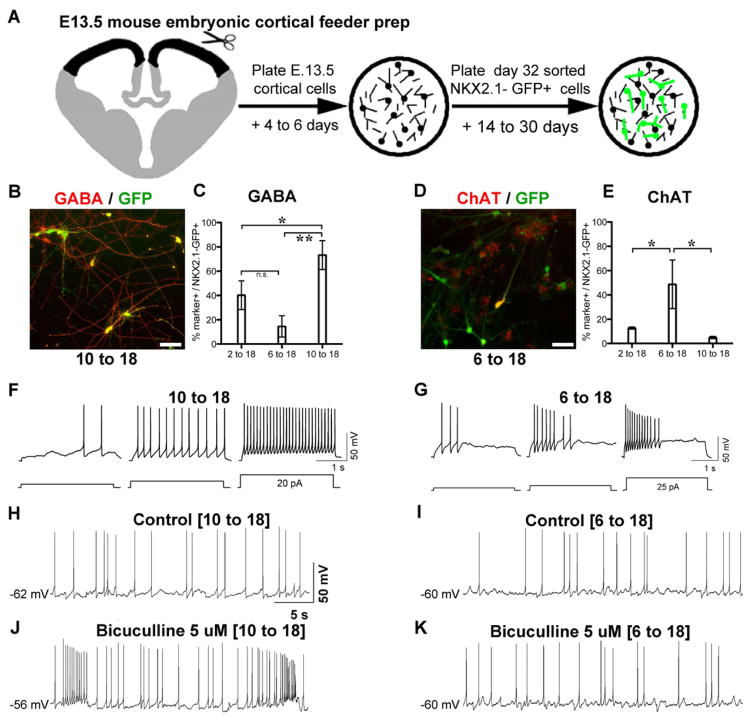
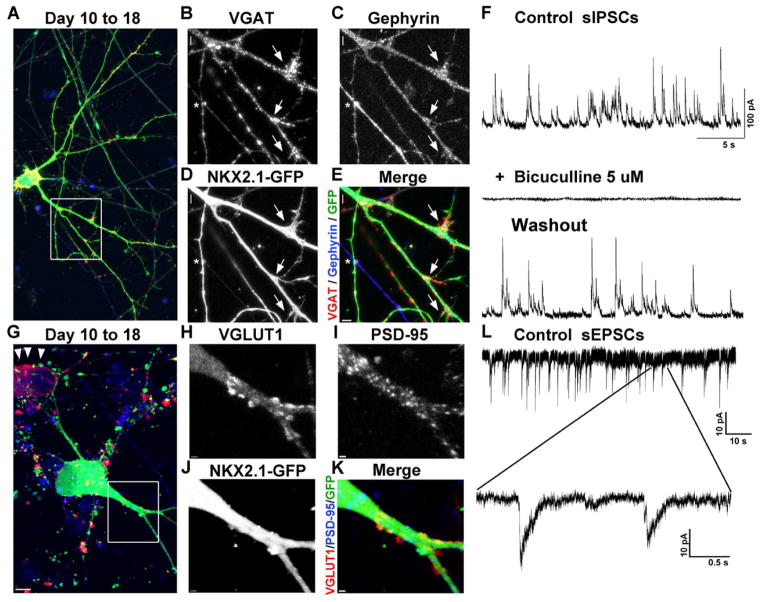
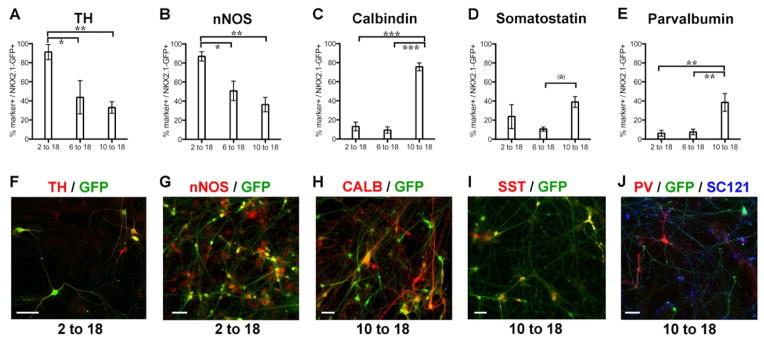
Comment in
-
Human cortical interneurons take their time.Cell Stem Cell. 2013 May 2;12(5):497-9. doi: 10.1016/j.stem.2013.04.017. Cell Stem Cell. 2013. PMID: 23642355
References
-
- Anderson SA, Classey JD, Conde F, Lund JS, Lewis DA. Synchronous development of pyramidal neuron dendritic spines and parvalbumin-immunoreactive chandelier neuron axon terminals in layer III of monkey prefrontal cortex. Neuroscience. 1995;67:7–22. - PubMed
-
- Anderson SA, Eisenstat DD, Shi L, Rubenstein JL. Interneuron migration from basal forebrain to neocortex: dependence on Dlx genes. Science. 1997;278:474–476. - PubMed
-
- Anderson SA, Marin O, Horn C, Jennings K, Rubenstein JL. Distinct cortical migrations from the medial and lateral ganglionic eminences. Development. 2001;128:353–363. - PubMed
-
- Baraban SC, Southwell DG, Estrada RC, Jones DL, Sebe JY, Alfaro-Cervello C, Garcia-Verdugo JM, Rubenstein JL, Alvarez-Buylla A. Reduction of seizures by transplantation of cortical GABAergic interneuron precursors into Kv1.1 mutant mice. Proceedings of the National Academy of Sciences of the United States of America. 2009;106:15472–15477. - PMC - PubMed
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
