

CTCF: the protein, the binding partners, the binding sites and their chromatin loops
- PMID: 23650640
- PMCID: PMC3682731
- DOI: 10.1098/rstb.2012.0369
CTCF: the protein, the binding partners, the binding sites and their chromatin loops
Abstract
CTCF has it all. The transcription factor binds to tens of thousands of genomic sites, some tissue-specific, others ultra-conserved. It can act as a transcriptional activator, repressor and insulator, and it can pause transcription. CTCF binds at chromatin domain boundaries, at enhancers and gene promoters, and inside gene bodies. It can attract many other transcription factors to chromatin, including tissue-specific transcriptional activators, repressors, cohesin and RNA polymerase II, and it forms chromatin loops. Yet, or perhaps therefore, CTCF's exact function at a given genomic site is unpredictable. It appears to be determined by the associated transcription factors, by the location of the binding site relative to the transcriptional start site of a gene, and by the site's engagement in chromatin loops with other CTCF-binding sites, enhancers or gene promoters. Here, we will discuss genome-wide features of CTCF binding events, as well as locus-specific functions of this remarkable transcription factor.
Keywords: CTCF; chromatin loops; cohesin; nuclear organization; transcription.
Figures
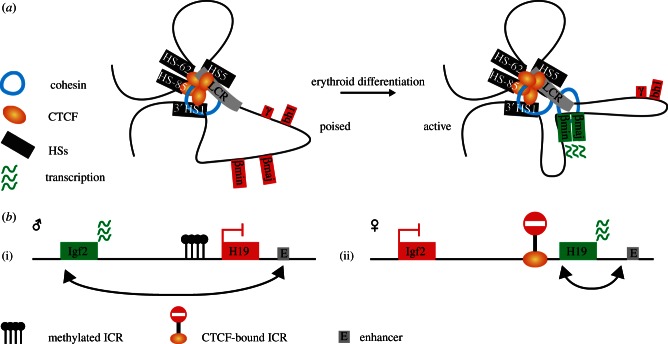
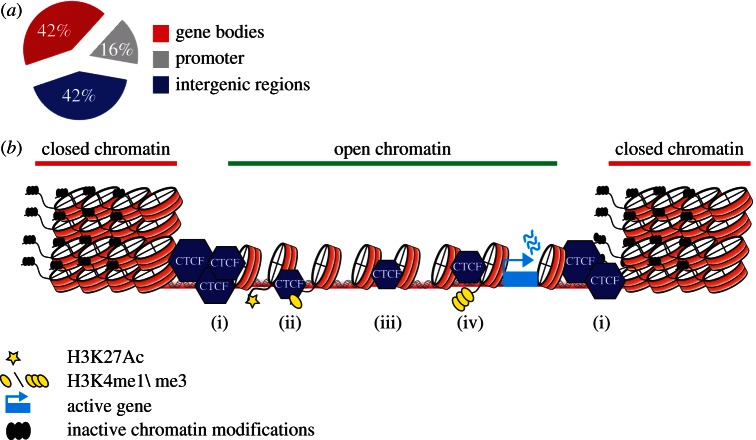
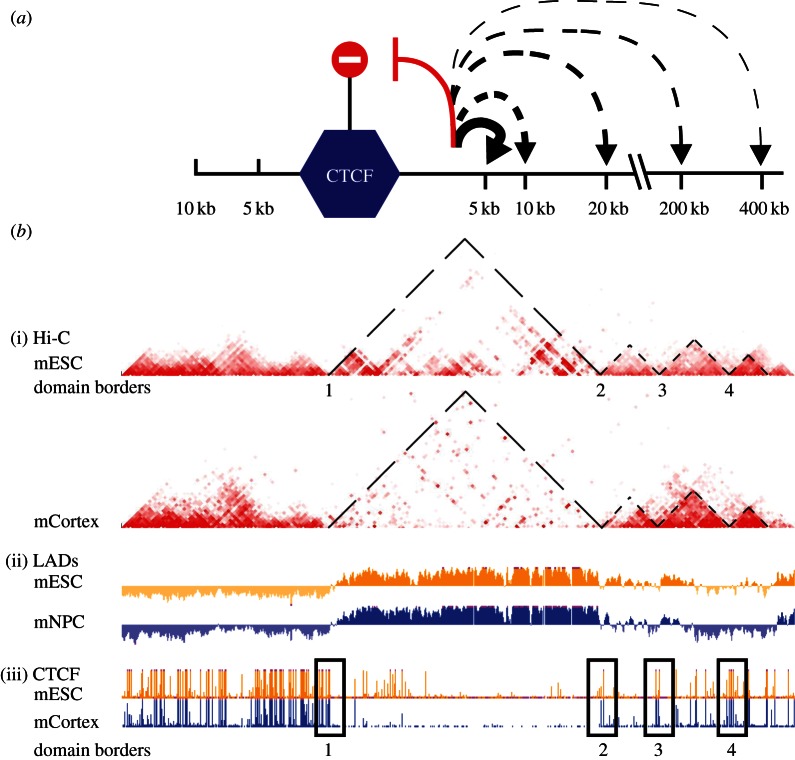
References
-
- Heath H, et al. 2008. CTCF regulates cell cycle progression of αβ T cells in the thymus. EMBO J. 27, 2839–2850 10.1038/emboj.2008.214 (doi:10.1038/emboj.2008.214) - DOI - PMC - PubMed
-
- Lobanenkov VV, Nicolas RH, Adler VV, Paterson H, Klenova EM, Polotskaja AV, Goodwin GH. 1990. A novel sequence-specific DNA binding protein which interacts with three regularly spaced direct repeats of the CCCTC-motif in the 5′-flanking sequence of the chicken c-myc gene. Oncogene 5, 1743–1753 - PubMed
-
- Klenova EM, Nicolas RH, Paterson HF, Carne AF, Heath CM, Goodwin GH, Neiman PE, Lobanenkov VV. 1993. CTCF, a conserved nuclear factor required for optimal transcriptional activity of the chicken c-myc gene, is an 11-Zn-finger protein differentially expressed in multiple forms. Mol. Cell. Biol. 13, 7612–7624 - PMC - PubMed
-
- Filippova GN, Fagerlie S, Klenova EM, Myers C, Dehner Y, Goodwin G, Neiman PE, Collins SJ, Lobanenkov VV. 1996. An exceptionally conserved transcriptional repressor, CTCF, employs different combinations of zinc fingers to bind diverged promoter sequences of avian and mammalian c-myc oncogenes. Mol. Cell. Biol. 16, 2802–2813 - PMC - PubMed
-
- Bell AC, West AG, Felsenfeld G. 1999. The protein CTCF is required for the enhancer blocking activity of vertebrate insulators. Cell 98, 387–396 10.1016/S0092-8674(00)81967-4 (doi:10.1016/S0092-8674(00)81967-4) - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous