

PTS phosphorylation of Mga modulates regulon expression and virulence in the group A streptococcus
- PMID: 23651410
- PMCID: PMC3702058
- DOI: 10.1111/mmi.12250
PTS phosphorylation of Mga modulates regulon expression and virulence in the group A streptococcus
Erratum in
- Mol Microbiol. 2014 Aug;93(4):834
Abstract
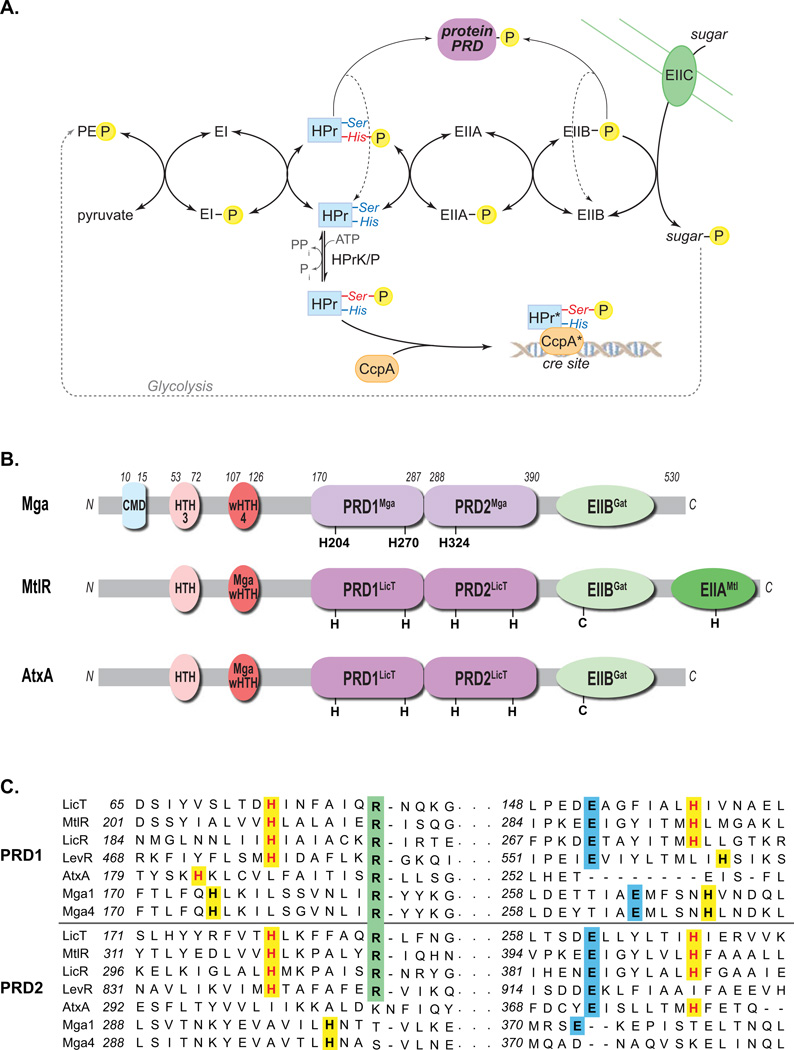
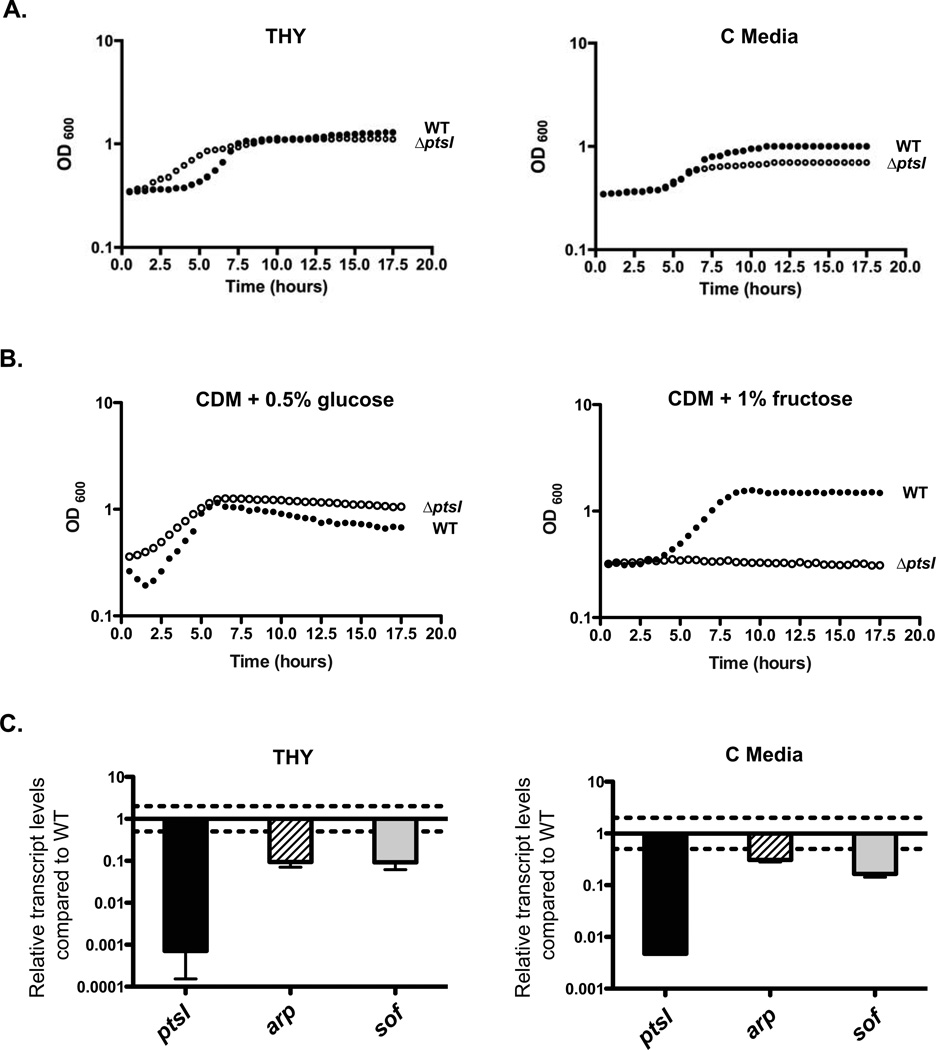
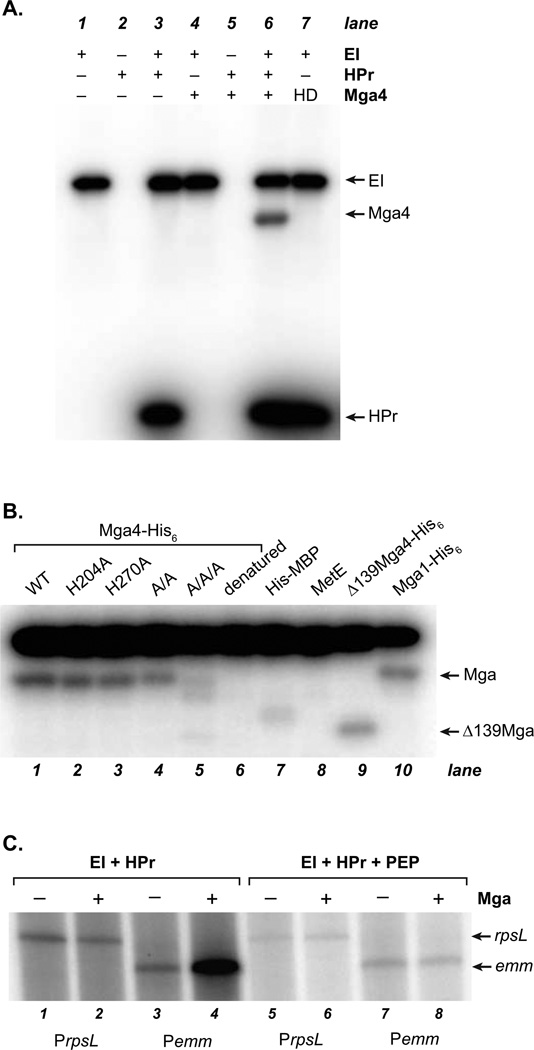
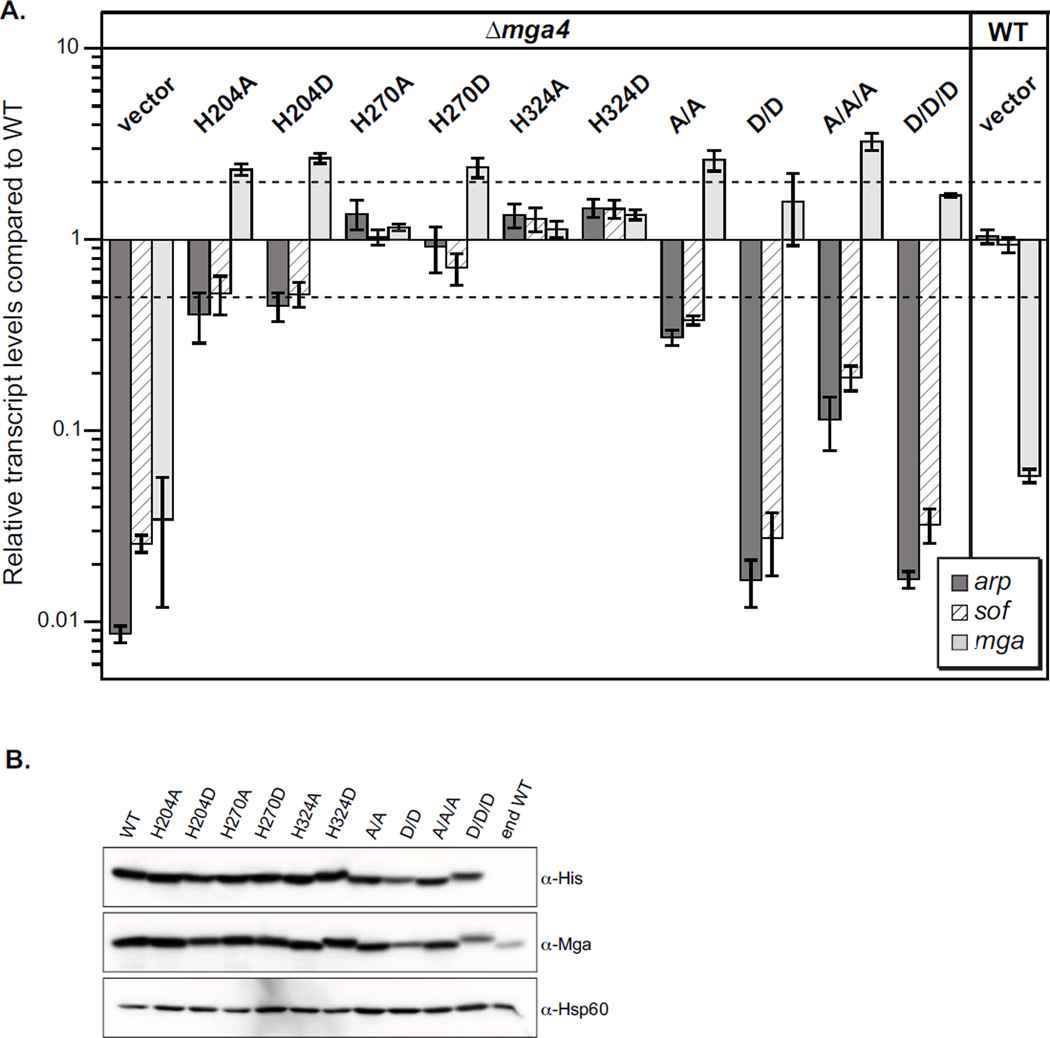
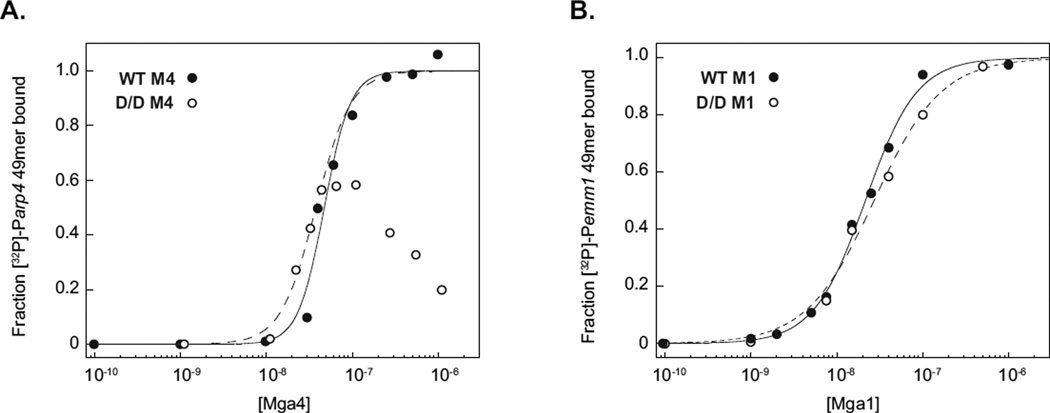
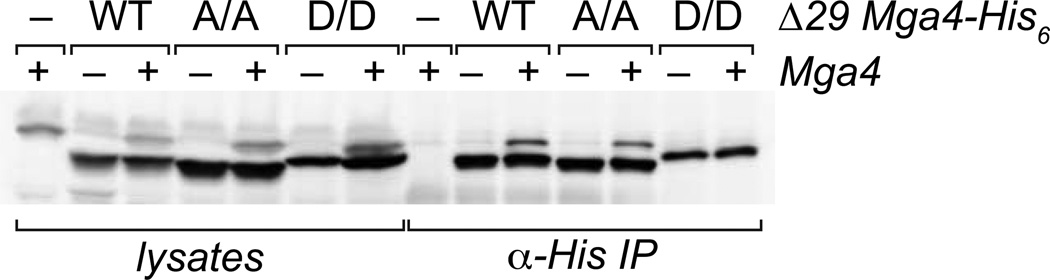
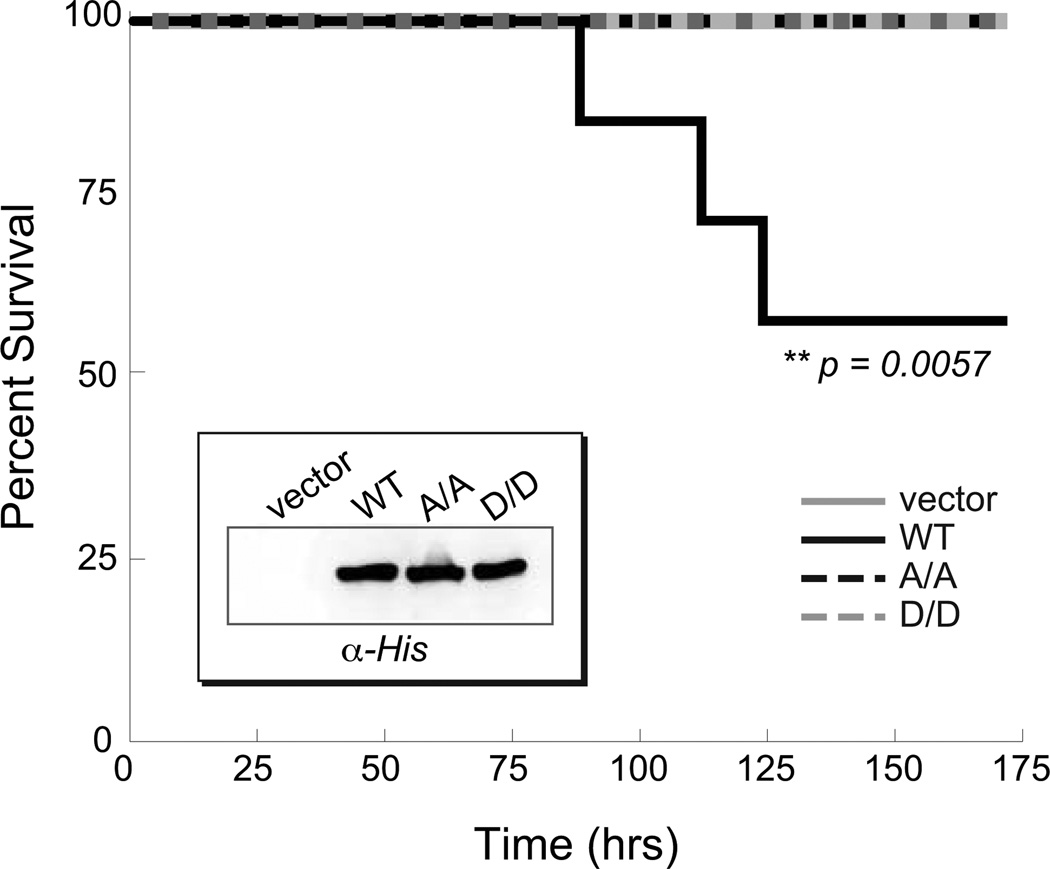
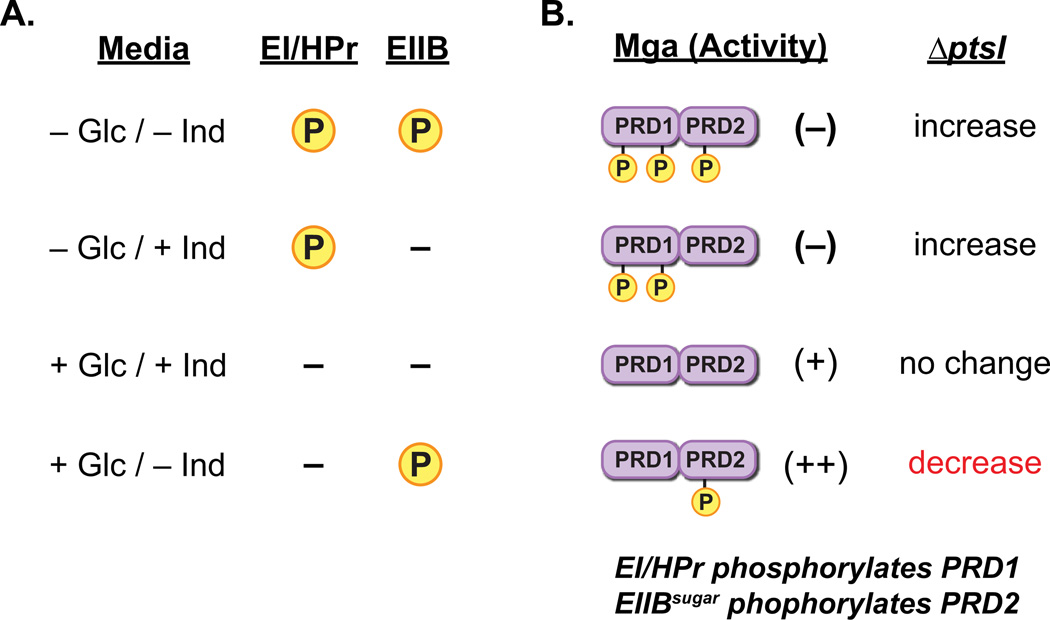
The ability of a bacterial pathogen to monitor available carbon sources in host tissues provides a clear fitness advantage. In the group A streptococcus (GAS), the virulence regulator Mga contains homology to phosphotransferase system (PTS) regulatory domains (PRDs) found in sugar operon regulators. Here we show that Mga was phosphorylated in vitro by the PTS components EI/HPr at conserved PRD histidines. A ΔptsI (EI-deficient) GAS mutant exhibited decreased Mga activity. However, PTS-mediated phosphorylation inhibited Mga-dependent transcription of emm in vitro. Using alanine (unphosphorylated) and aspartate (phosphomimetic) mutations of PRD histidines, we establish that a doubly phosphorylated PRD1 phosphomimetic (D/DMga4) is completely inactive in vivo, shutting down expression of the Mga regulon. Although D/DMga4 is still able to bind DNA in vitro, homo-multimerization of Mga is disrupted and the protein is unable to activate transcription. PTS-mediated regulation of Mga activity appears to be important for pathogenesis, as bacteria expressing either non-phosphorylated (A/A) or phosphomimetic (D/D) PRD1 Mga mutants were attenuated in a model of GAS invasive skin disease. Thus, PTS-mediated phosphorylation of Mga may allow the bacteria to modulate virulence gene expression in response to carbohydrate status. Furthermore, PRD-containing virulence regulators (PCVRs) appear to be widespread in Gram-positive pathogens.
© 2013 John Wiley & Sons Ltd.
Figures
Similar articles
-
The Streptococcus pyogenes stand-alone regulator RofA exhibits characteristics of a PRD-containing virulence regulator.Infect Immun. 2024 Jun 11;92(6):e0008324. doi: 10.1128/iai.00083-24. Epub 2024 May 7. Infect Immun. 2024. PMID: 38712951 Free PMC article.
-
Glucose Levels Alter the Mga Virulence Regulon in the Group A Streptococcus.Sci Rep. 2018 Mar 21;8(1):4971. doi: 10.1038/s41598-018-23366-7. Sci Rep. 2018. PMID: 29563558 Free PMC article.
-
Phosphorylation events in the multiple gene regulator of group A Streptococcus significantly influence global gene expression and virulence.Infect Immun. 2015 Jun;83(6):2382-95. doi: 10.1128/IAI.03023-14. Epub 2015 Mar 30. Infect Immun. 2015. PMID: 25824840 Free PMC article.
-
The Mga virulence regulon: infection where the grass is greener.Mol Microbiol. 2007 Dec;66(5):1056-65. doi: 10.1111/j.1365-2958.2007.06006.x. Mol Microbiol. 2007. PMID: 18001346 Review.
-
PRD-Containing Virulence Regulators (PCVRs) in Pathogenic Bacteria.Front Cell Infect Microbiol. 2021 Oct 19;11:772874. doi: 10.3389/fcimb.2021.772874. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 34737980 Free PMC article. Review.
Cited by
-
Glucose Phosphotransferase System Modulates Pyruvate Metabolism, Bacterial Fitness, and Microbial Ecology in Oral Streptococci.J Bacteriol. 2023 Jan 26;205(1):e0035222. doi: 10.1128/jb.00352-22. Epub 2022 Dec 5. J Bacteriol. 2023. PMID: 36468868 Free PMC article.
-
A Conserved Streptococcal Virulence Regulator Controls the Expression of a Distinct Class of M-Like Proteins.mBio. 2019 Oct 22;10(5):e02500-19. doi: 10.1128/mBio.02500-19. mBio. 2019. PMID: 31641092 Free PMC article.
-
Enterococcus faecalis Gluconate Phosphotransferase System Accelerates Experimental Colitis and Bacterial Killing by Macrophages.Infect Immun. 2019 Jun 20;87(7):e00080-19. doi: 10.1128/IAI.00080-19. Print 2019 Jul. Infect Immun. 2019. PMID: 31036600 Free PMC article.
-
The bacterial phosphoenolpyruvate:carbohydrate phosphotransferase system: regulation by protein phosphorylation and phosphorylation-dependent protein-protein interactions.Microbiol Mol Biol Rev. 2014 Jun;78(2):231-56. doi: 10.1128/MMBR.00001-14. Microbiol Mol Biol Rev. 2014. PMID: 24847021 Free PMC article. Review.
-
Streptococcus pyogenes polymyxin B-resistant mutants display enhanced ExPortal integrity.J Bacteriol. 2014 Jul;196(14):2563-77. doi: 10.1128/JB.01596-14. Epub 2014 May 2. J Bacteriol. 2014. PMID: 24794568 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
