

A network of substrates of the E3 ubiquitin ligases MDM2 and HUWE1 control apoptosis independently of p53
- PMID: 23652204
- PMCID: PMC3770270
- DOI: 10.1126/scisignal.2003741
A network of substrates of the E3 ubiquitin ligases MDM2 and HUWE1 control apoptosis independently of p53
Abstract
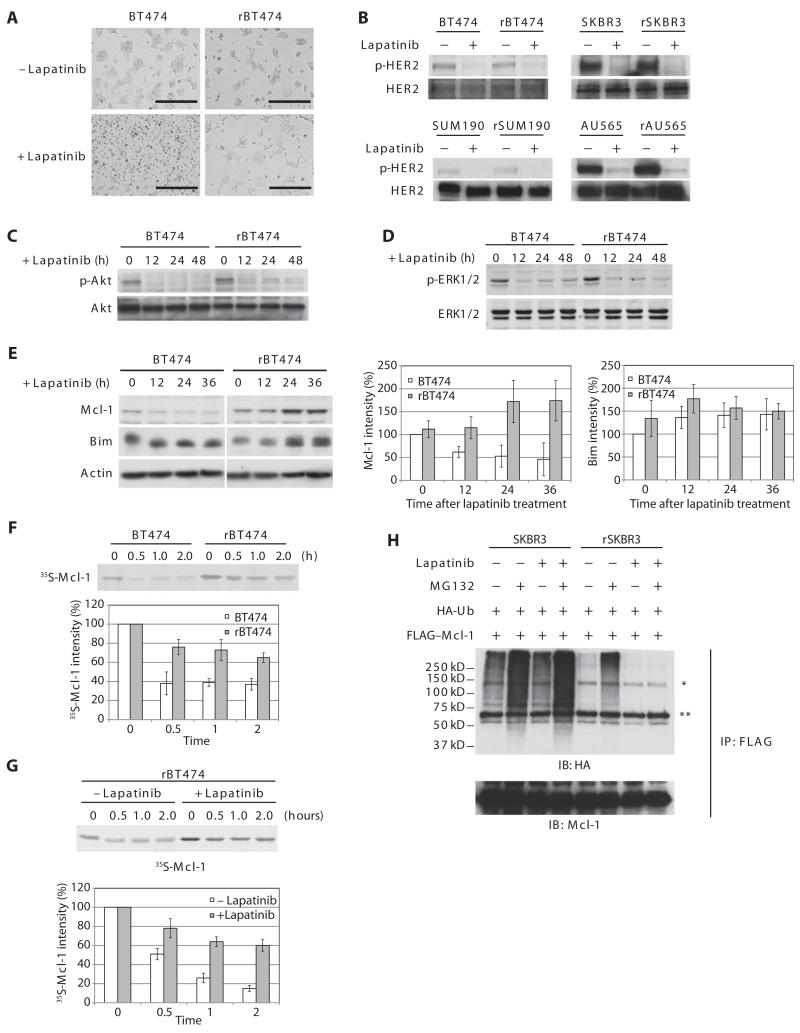
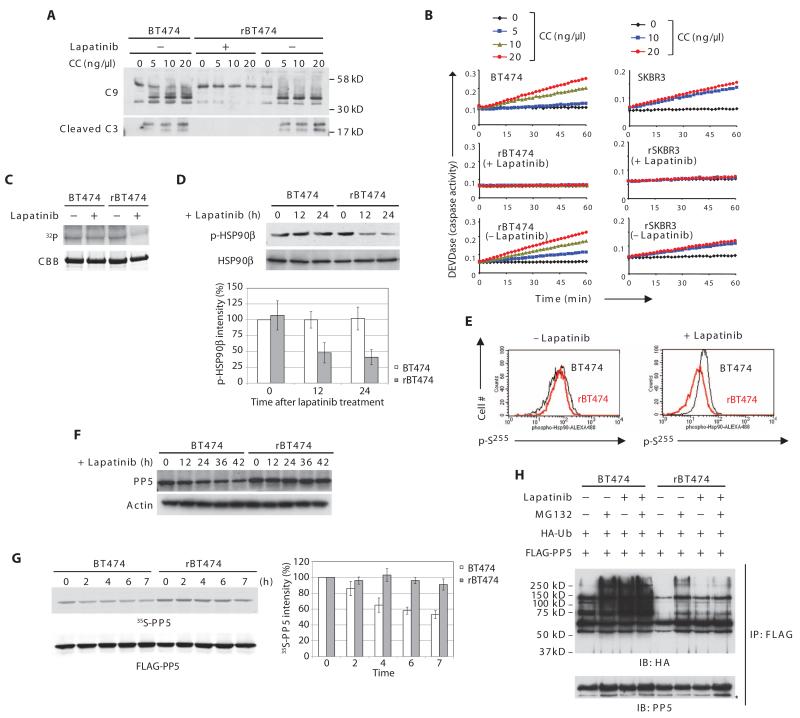
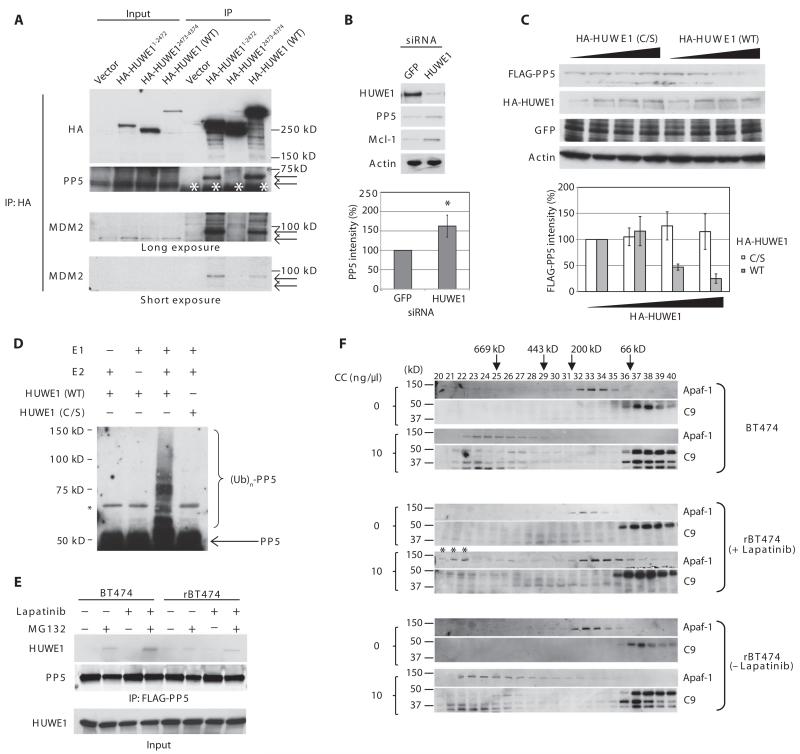
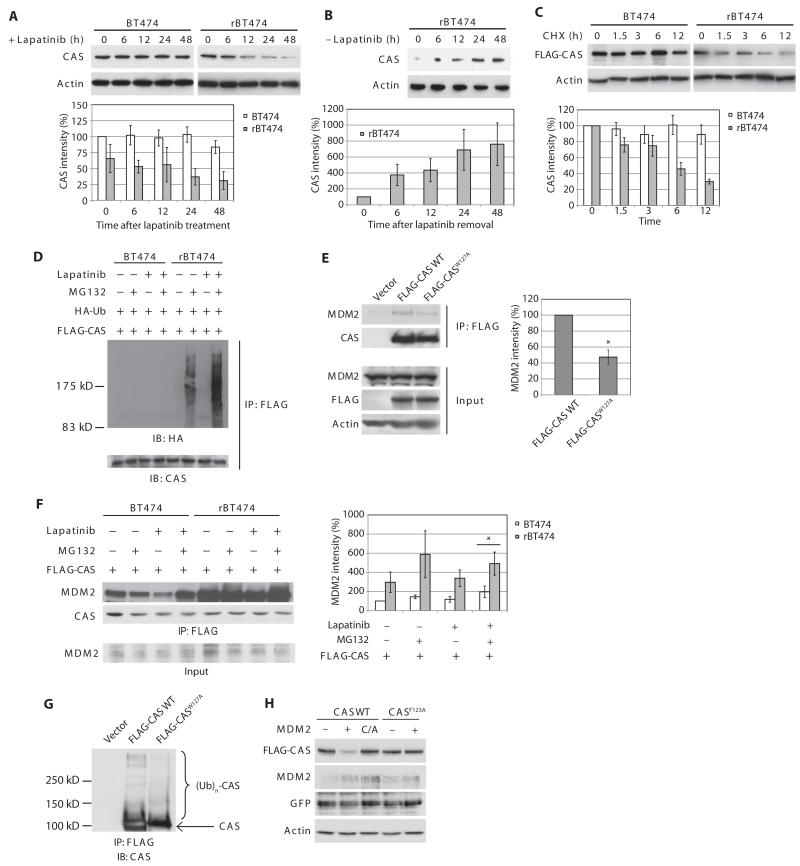
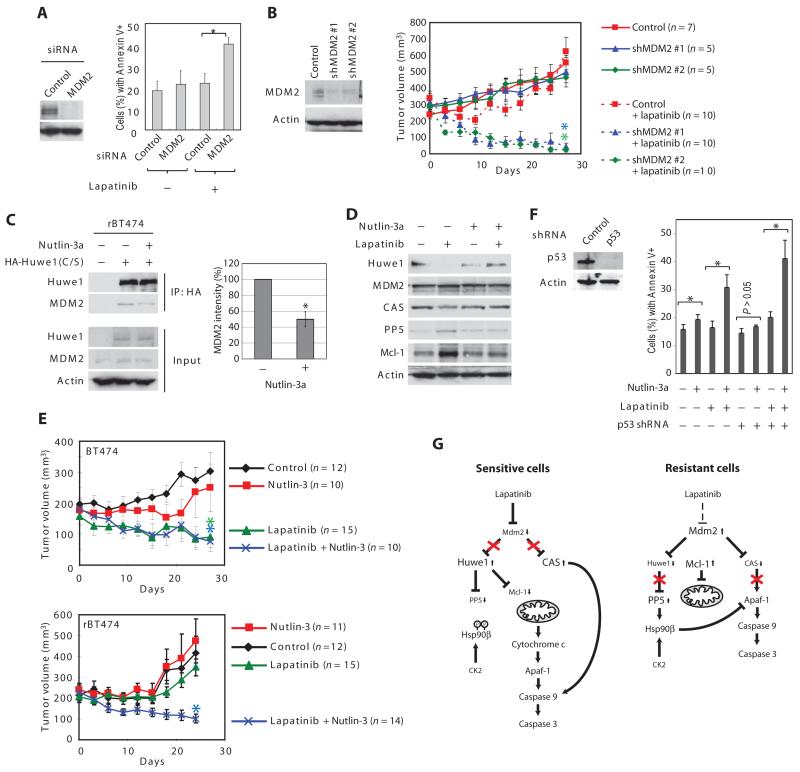
In the intrinsic pathway of apoptosis, cell-damaging signals promote the release of cytochrome c from mitochondria, triggering activation of the Apaf-1 and caspase-9 apoptosome. The ubiquitin E3 ligase MDM2 decreases the stability of the proapoptotic factor p53. We show that it also coordinated apoptotic events in a p53-independent manner by ubiquitylating the apoptosome activator CAS and the ubiquitin E3 ligase HUWE1. HUWE1 ubiquitylates the antiapoptotic factor Mcl-1, and we found that HUWE1 also ubiquitylated PP5 (protein phosphatase 5), which indirectly inhibited apoptosome activation. Breast cancers that are positive for the tyrosine receptor kinase HER2 (human epidermal growth factor receptor 2) tend to be highly aggressive. In HER2-positive breast cancer cells treated with the HER2 tyrosine kinase inhibitor lapatinib, MDM2 was degraded and HUWE1 was stabilized. In contrast, in breast cancer cells that acquired resistance to lapatinib, the abundance of MDM2 was not decreased and HUWE1 was degraded, which inhibited apoptosis, regardless of p53 status. MDM2 inhibition overcame lapatinib resistance in cells with either wild-type or mutant p53 and in xenograft models. These findings demonstrate broader, p53-independent roles for MDM2 and HUWE1 in apoptosis and specifically suggest the potential for therapy directed against MDM2 to overcome lapatinib resistance.
Figures

References
-
- Hollstein M, Sidransky D, Vogelstein B, Harris CC. p53 mutations in human cancers. Science. 1991;253:49–53. - PubMed
-
- Feki A, Irminger-Finger I. Mutational spectrum of p53 mutations in primary breast and ovarian tumors. Crit. Rev. Oncol. Hematol. 2004;52:103–116. - PubMed
-
- Momand J, Zambetti GP, Olson DC, George D, Levine AJ. The mdm-2 oncogene product forms a complex with the p53 protein and inhibits p53-mediated transactivation. Cell. 1992;69:1237–1245. - PubMed
-
- Oliner JD, Pietenpol JA, Thiagalingam S, Gyuris J, Kinzler KW, Vogelstein B. Oncoprotein MDM2 conceals the activation domain of tumour suppressor p53. Nature. 1993;362:857–860. - PubMed
-
- Haupt Y, Maya R, Kazaz A, Oren M. Mdm2 promotes the rapid degradation of p53. Nature. 1997;387:296–299. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
