

The genetic basis of speciation in the Giliopsis lineage of Ipomopsis (Polemoniaceae)
- PMID: 23652565
- PMCID: PMC3746823
- DOI: 10.1038/hdy.2013.41
The genetic basis of speciation in the Giliopsis lineage of Ipomopsis (Polemoniaceae)
Abstract
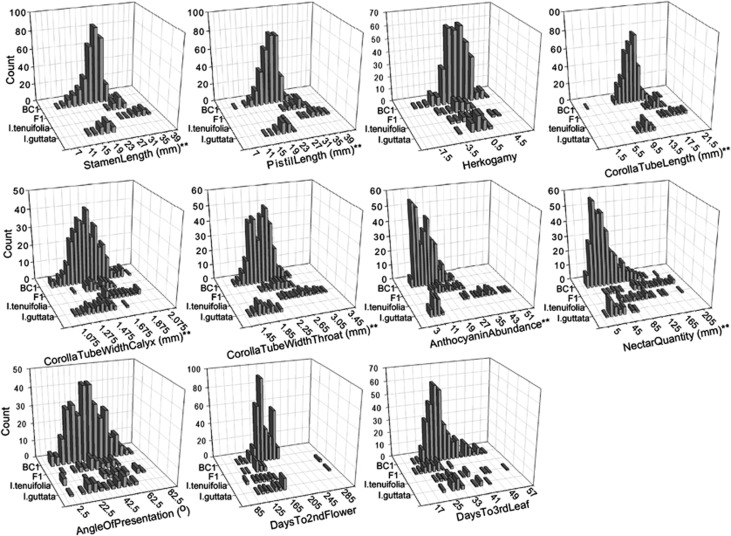
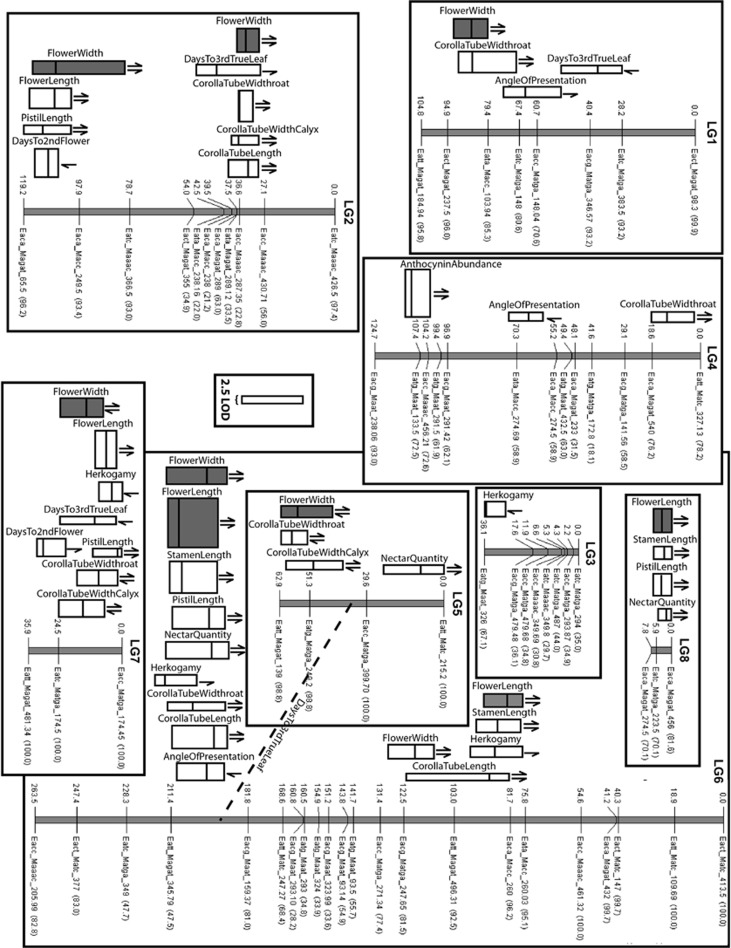
One of the most powerful drivers of speciation in plants is pollinator-mediated disruptive selection, which leads to the divergence of floral traits adapted to the morphology and behavior of different pollinators. Despite the widespread importance of this speciation mechanism, its genetic basis has been explored in only a few groups. Here, we characterize the genetic basis of pollinator-mediated divergence of two species in genus Ipomopsis, I. guttata and I. tenuifolia, using quantitative trait locus (QTL) analyses of floral traits and other variable phenotypes. We detected one to six QTLs per trait, with each QTL generally explaining small to modest amounts of the phenotypic variance of a backcross hybrid population. In contrast, flowering time and anthocyanin abundance (a metric of color variation) were controlled by a few QTLs of relatively large effect. QTLs were strongly clustered within linkage groups, with 26 of 37 QTLs localized to six marker-interval 'hotspots,' all of which harbored pleiotropic QTLs. In contrast to other studies that have examined the genetic basis of pollinator shifts, our results indicate that, in general, mutations of small to modest effect on phenotype were involved. Thus, the evolutionary transition between the distinct pollination modes of I. guttata and I. tenuifolia likely proceeded incrementally, rather than saltationally.
Figures

Similar articles
-
Genetic Mapping Reveals an Anthocyanin Biosynthesis Pathway Gene Potentially Influencing Evolutionary Divergence between Two Subspecies of Scarlet Gilia (Ipomopsis aggregata).Mol Biol Evol. 2018 Apr 1;35(4):807-822. doi: 10.1093/molbev/msx318. Mol Biol Evol. 2018. PMID: 29253197
-
Adaptive differentiation in floral traits in the presence of high gene flow in scarlet gilia (Ipomopsis aggregata).Mol Ecol. 2016 Dec;25(23):5862-5875. doi: 10.1111/mec.13752. Epub 2016 Aug 8. Mol Ecol. 2016. PMID: 27392816
-
Complex interactions underlie the correlated evolution of floral traits and their association with pollinators in a clade with diverse pollination systems.Evolution. 2021 Jun;75(6):1431-1449. doi: 10.1111/evo.14220. Epub 2021 Apr 16. Evolution. 2021. PMID: 33818785
-
Pleiotropy and the evolution of floral integration.New Phytol. 2016 Jan;209(1):80-5. doi: 10.1111/nph.13583. Epub 2015 Jul 30. New Phytol. 2016. PMID: 26224529 Review.
-
The genetic control of flower-pollinator specificity.Curr Opin Plant Biol. 2013 Aug;16(4):422-8. doi: 10.1016/j.pbi.2013.05.004. Epub 2013 Jun 10. Curr Opin Plant Biol. 2013. PMID: 23763819 Free PMC article. Review.
Cited by
-
The genetics of reproductive organ morphology in two Petunia species with contrasting pollination syndromes.Planta. 2015 May;241(5):1241-54. doi: 10.1007/s00425-015-2251-2. Epub 2015 Feb 6. Planta. 2015. PMID: 25656052
-
Genomics of plant speciation.Plant Commun. 2023 Sep 11;4(5):100599. doi: 10.1016/j.xplc.2023.100599. Epub 2023 Apr 11. Plant Commun. 2023. PMID: 37050879 Free PMC article. Review.
-
Genetic architecture of pollination syndrome transition between hummingbird-specialist and generalist species in the genus Rhytidophyllum (Gesneriaceae).PeerJ. 2015 Jun 18;3:e1028. doi: 10.7717/peerj.1028. eCollection 2015. PeerJ. 2015. PMID: 26157613 Free PMC article.
-
Geometric morphometrics reveals shifts in flower shape symmetry and size following gene knockdown of CYCLOIDEA and ANTHOCYANIDIN SYNTHASE.BMC Plant Biol. 2017 Nov 17;17(1):205. doi: 10.1186/s12870-017-1152-x. BMC Plant Biol. 2017. PMID: 29149840 Free PMC article.
-
Genetic architecture of a pollinator shift and its fate in secondary hybrid zones of two Petunia species.BMC Biol. 2023 Mar 20;21(1):58. doi: 10.1186/s12915-023-01561-x. BMC Biol. 2023. PMID: 36941631 Free PMC article.
References
-
- Beavis WD.1998QTL analyses: power, precision, and accuracyIn: Paterson AH, (ed.)Molecular Dissection of Complex Traits CRC Press: Boca Raton, FL; 145–162.
-
- Bishop DT, Cannings C, Skolnick M, Williamson JA.1983The number of polymorphic clones required to map the human genomeIn: Weir BS, (ed.)Statistical Analysis of DNA Sequence Data. M Dekker: New York; 181–200.
-
- Bleiweiss R. Mimicry on the QT(L): Genetics of speciation in Mimulus. Evolution. 2001;55:1706–1709. - PubMed
-
- Bolten AB, Feinsinger P. Why Do Hummingbird Flowers Secrete Dilute Nectar. Biotropica. 1978;10:307–309.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
