

Structural analysis of collagen type I interactions with human fibronectin reveals a cooperative binding mode
- PMID: 23653354
- PMCID: PMC3682544
- DOI: 10.1074/jbc.M113.469841
Structural analysis of collagen type I interactions with human fibronectin reveals a cooperative binding mode
Abstract
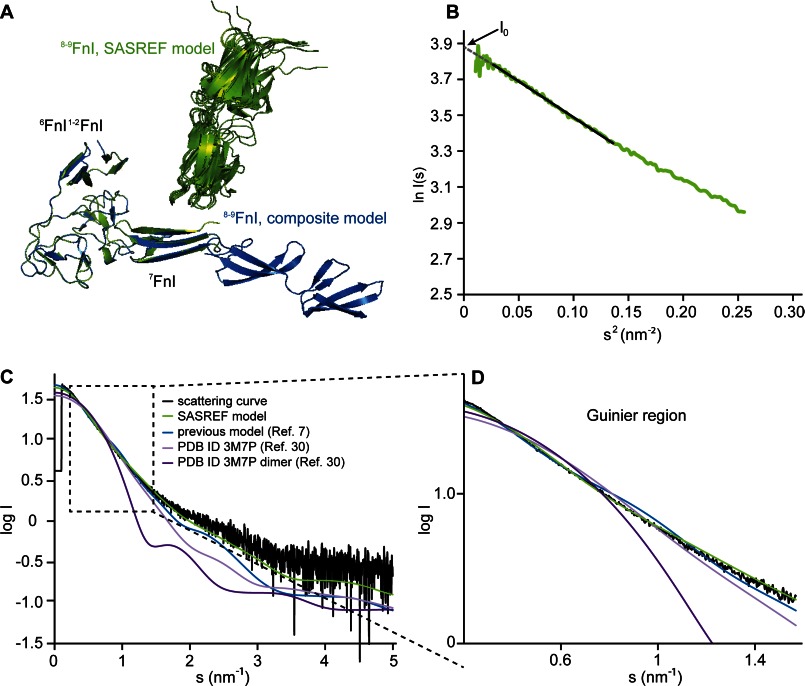
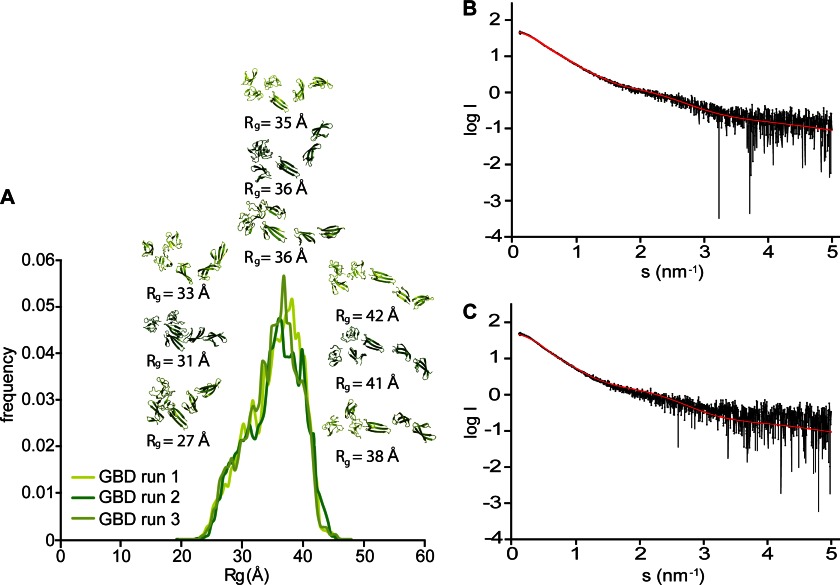
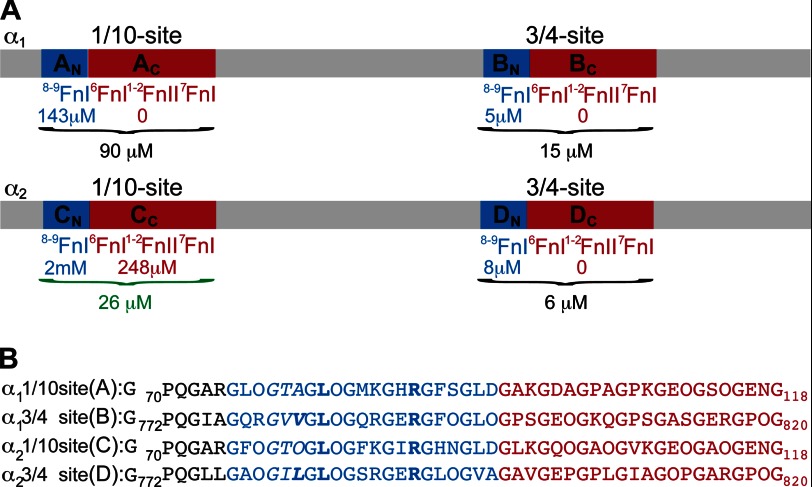
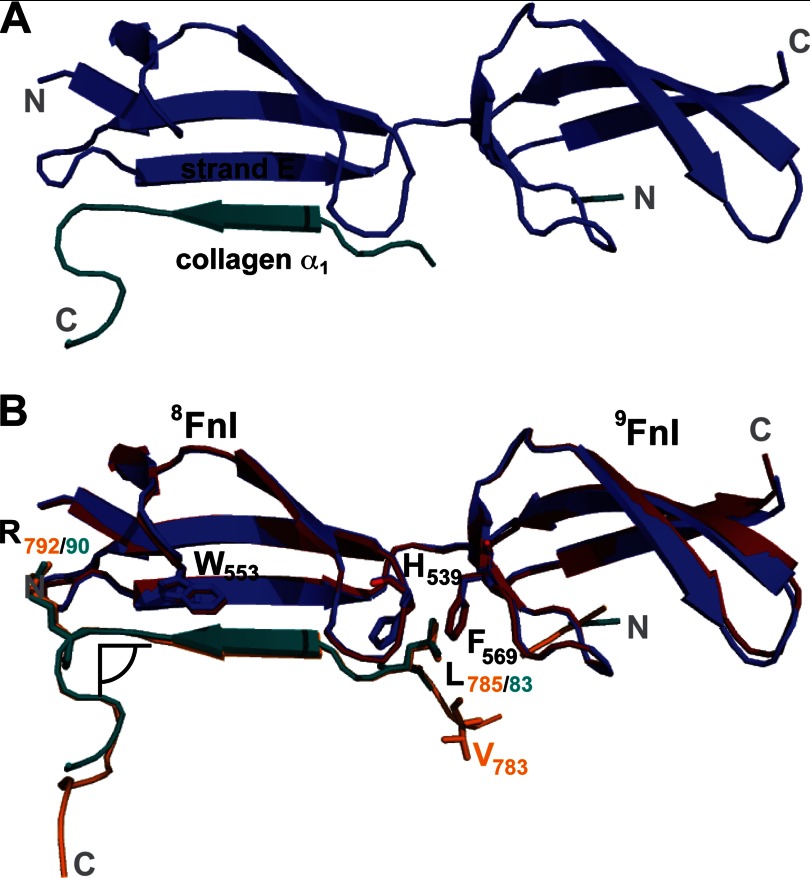
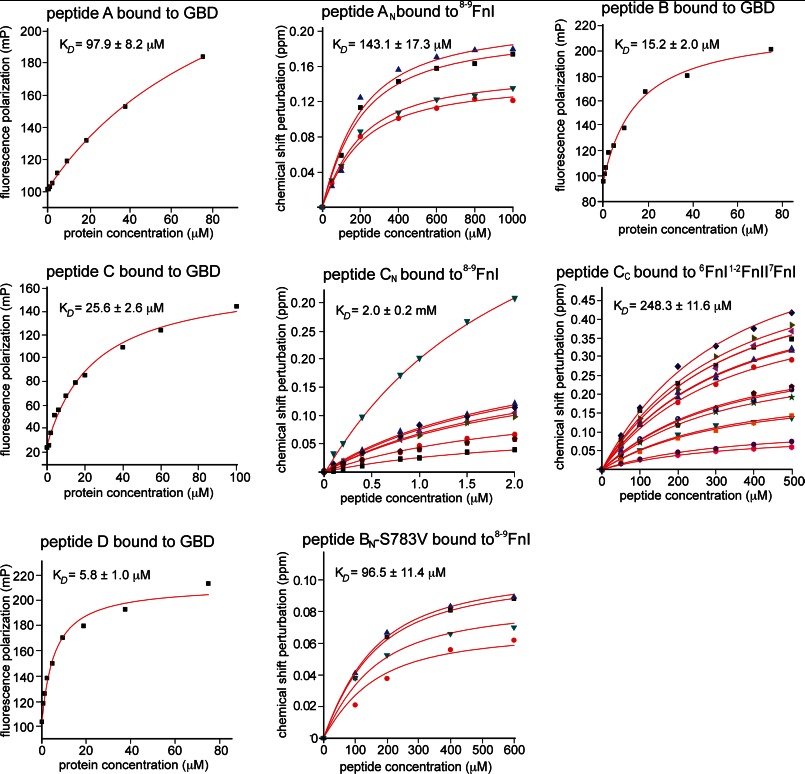
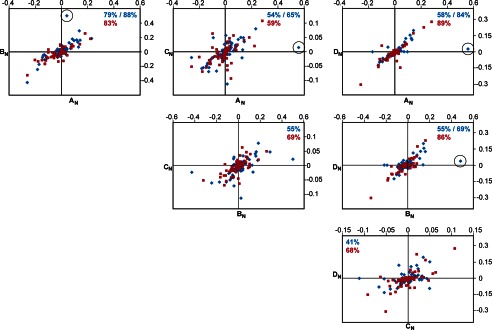
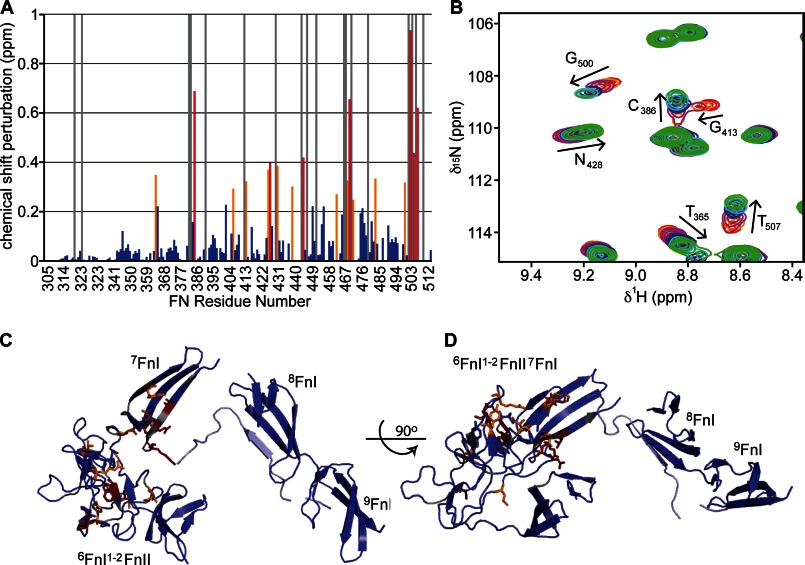
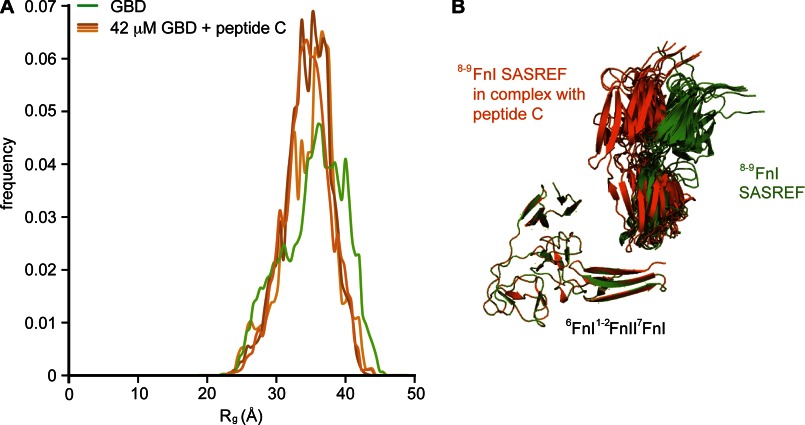
Despite its biological importance, the interaction between fibronectin (FN) and collagen, two abundant and crucial tissue components, has not been well characterized on a structural level. Here, we analyzed the four interactions formed between epitopes of collagen type I and the collagen-binding fragment (gelatin-binding domain (GBD)) of human FN using solution NMR, fluorescence, and small angle x-ray scattering methods. Collagen association with FN modules (8-9)FnI occurs through a conserved structural mechanism but exhibits a 400-fold disparity in affinity between collagen sites. This disparity is reduced in the full-length GBD, as (6)FnI(1-2)FnII(7)FnI binds a specific collagen epitope next to the weakest (8-9)FnI-binding site. The cooperative engagement of all GBD modules with collagen results in four broadly equipotent FN-collagen interaction sites. Collagen association stabilizes a distinct monomeric GBD conformation in solution, giving further evidence to the view that FN fragments form well defined functional and structural units.
Keywords: Collagen; Extracellular Matrix Proteins; Fibronectin; GBD; NMR; SAXS; X-ray Crystallography.
Figures
References
-
- George E. L., Georges-Labouesse E. N., Patel-King R. S., Rayburn H., Hynes R. O. (1993) Defects in mesoderm, neural tube and vascular development in mouse embryos lacking fibronectin. Development 119, 1079–1091 - PubMed
-
- Leitinger B., Hohenester E. (2007) Mammalian collagen receptors. Matrix Biol. 26, 146–155 - PubMed
-
- Campbell I. D., Downing A. K. (1998) NMR of modular proteins. Nat. Struct. Biol. 5, 496–499 - PubMed
-
- Leahy D. J., Aukhil I., Erickson H. P. (1996) 2.0 Å crystal structure of a four-domain segment of human fibronectin encompassing the RGD loop and synergy region. Cell 84, 155–164 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
