

Latency of tone-burst-evoked auditory brain stem responses and otoacoustic emissions: level, frequency, and rise-time effects
- PMID: 23654387
- PMCID: PMC3663861
- DOI: 10.1121/1.4798666
Latency of tone-burst-evoked auditory brain stem responses and otoacoustic emissions: level, frequency, and rise-time effects
Abstract
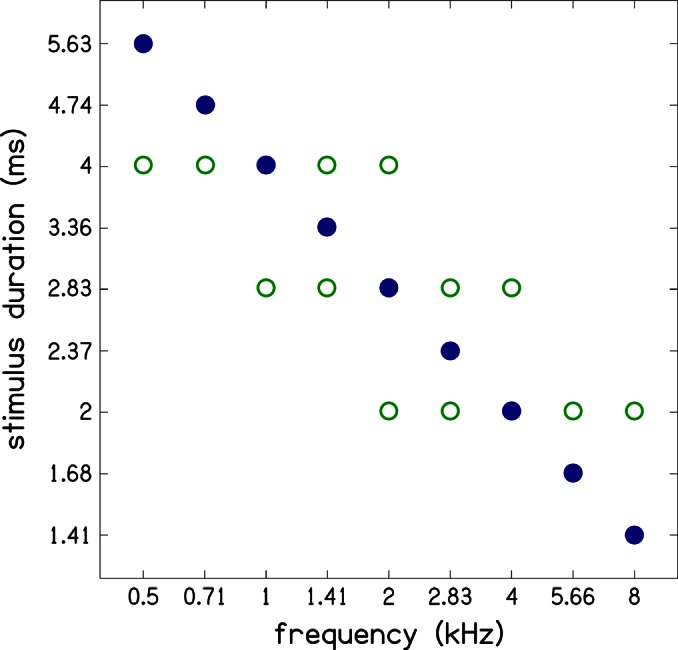
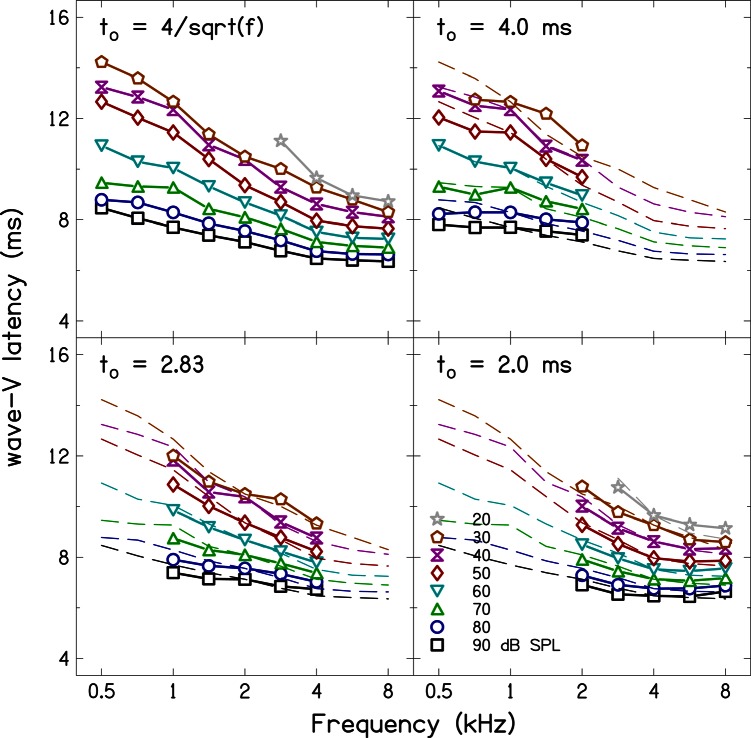
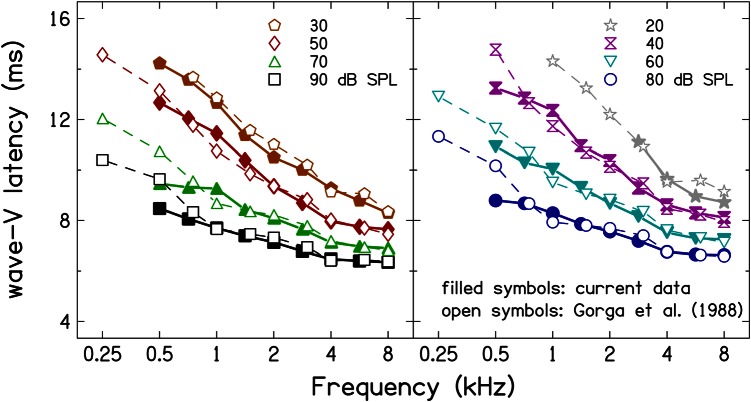
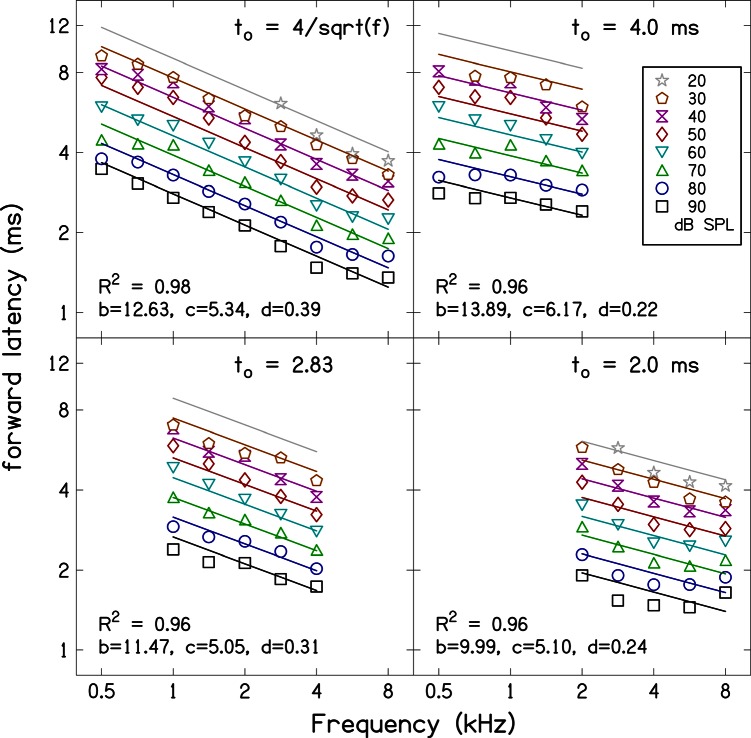
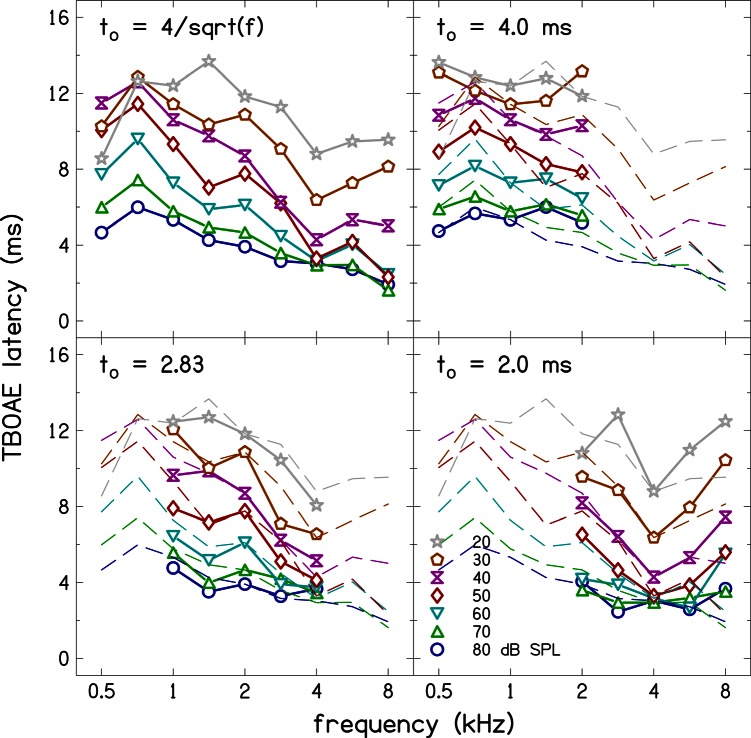
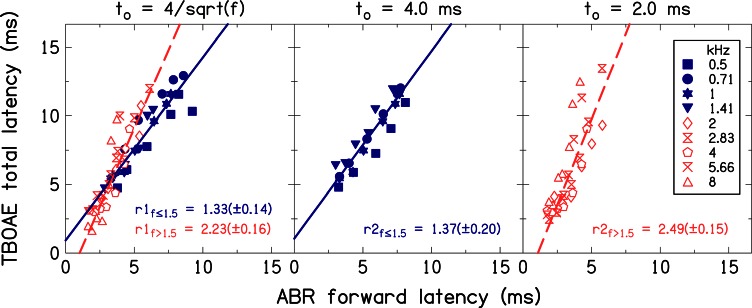
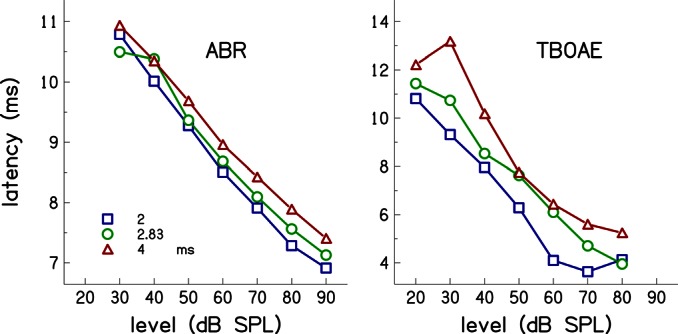
Simultaneous measurement of auditory brain stem response (ABR) and otoacoustic emission (OAE) delays may provide insights into effects of level, frequency, and stimulus rise-time on cochlear delay. Tone-burst-evoked ABRs and OAEs (TBOAEs) were measured simultaneously in normal-hearing human subjects. Stimuli included a wide range of frequencies (0.5-8 kHz), levels (20-90 dB SPL), and tone-burst rise times. ABR latencies have orderly dependence on these three parameters, similar to previously reported data by Gorga et al. [J. Speech Hear. Res. 31, 87-97 (1988)]. Level dependence of ABR and TBOAE latencies was similar across a wide range of stimulus conditions. At mid-frequencies, frequency dependence of ABR and TBOAE latencies were similar. The dependence of ABR latency on both rise time and level was significant; however, the interaction was not significant, suggesting independent effects. Comparison between ABR and TBOAE latencies reveals that the ratio of TBOAE latency to ABR forward latency (the level-dependent component of ABR total latency) is close to one below 1.5 kHz, but greater than two above 1.5 kHz. Despite the fact that the current experiment was designed to test compatibility with models of reverse-wave propagation, existing models do not completely explain the current data.
Figures


References
-
- Burkard, R., and Secor, C. (2002). “ Overview of auditory evoked potential,” in Handbook of Clinical Audiology, edited by Katz J. (Lippincott, Williams, and Wilkins, Philadelphia, PA: ), Chap. 14, pp. 233–248.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous