

Transcription factor-based screens and synthetic selections for microbial small-molecule biosynthesis
- PMID: 23656325
- PMCID: PMC11245165
- DOI: 10.1021/sb300091d
Transcription factor-based screens and synthetic selections for microbial small-molecule biosynthesis
Abstract
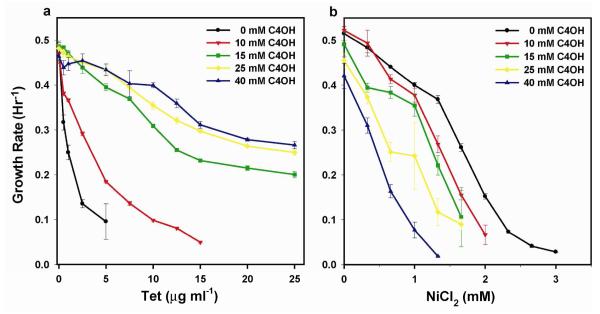
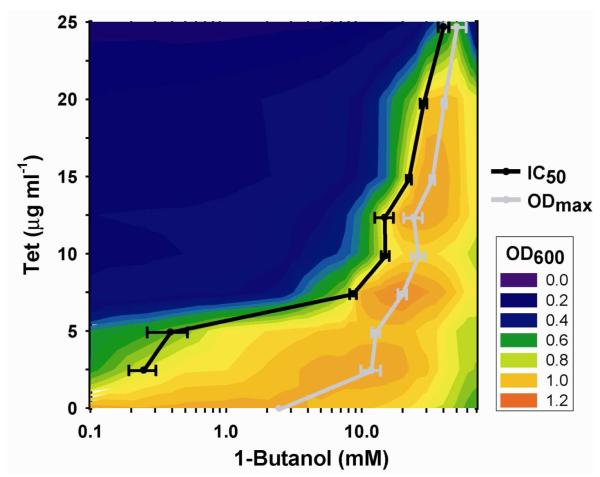
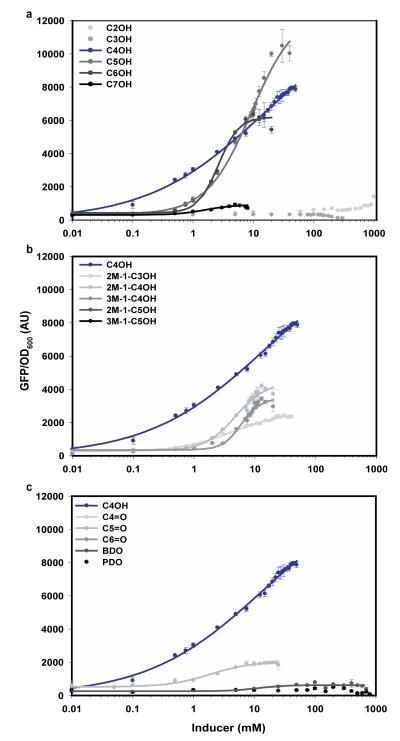
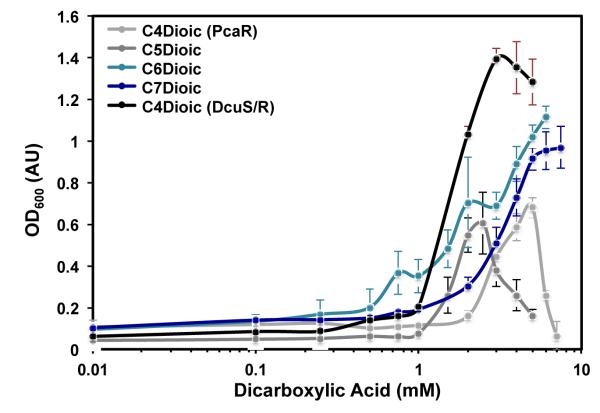
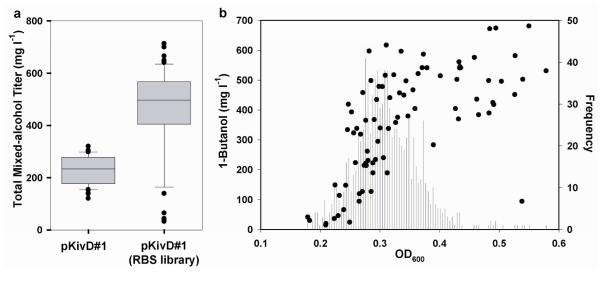
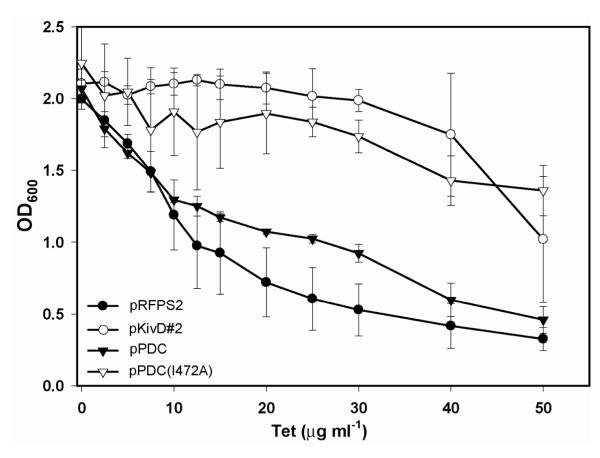
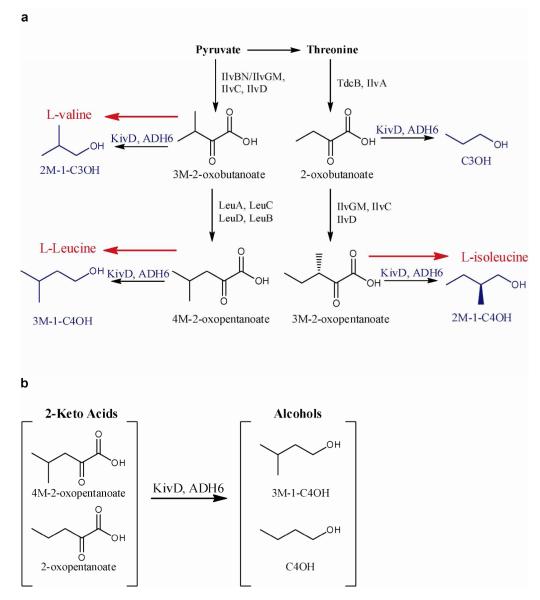
Continued advances in metabolic engineering are increasing the number of small molecules being targeted for microbial production. Pathway yields and productivities, however, are often suboptimal, and strain improvement remains a persistent challenge given that the majority of small molecules are difficult to screen for and their biosynthesis does not improve host fitness. In this work, we have developed a generalized approach to screen or select for improved small-molecule biosynthesis using transcription factor-based biosensors. Using a tetracycline resistance gene 3' of a small-molecule inducible promoter, host antibiotic resistance, and hence growth rate, was coupled to either small-molecule concentration in the growth medium or a small-molecule production phenotype. Biosensors were constructed for two important chemical classes, dicarboxylic acids and alcohols, using transcription factor-promoter pairs derived from Pseudomonas putida, Thauera butanivorans, or E. coli. Transcription factors were selected for specific activation by either succinate, adipate, or 1-butanol, and we demonstrate product-dependent growth in E. coli using all three compounds. The 1-butanol biosensor was applied in a proof-of-principle liquid culture screen to optimize 1-butanol biosynthesis in engineered E. coli, identifying a pathway variant yielding a 35% increase in 1-butanol specific productivity through optimization of enzyme expression levels. Lastly, to demonstrate the capacity to select for enzymatic activity, the 1-butanol biosensor was applied as synthetic selection, coupling in vivo 1-butanol biosynthesis to E. coli fitness, and an 120-fold enrichment for a 1-butanol production phenotype was observed following a single round of positive selection.
Figures
References
-
- Keasling JD. Manufacturing molecules through metabolic engineering. Science. 2010;330:1355–1358. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
