

Insights into β-lactamases from Burkholderia species, two phylogenetically related yet distinct resistance determinants
- PMID: 23658015
- PMCID: PMC3696682
- DOI: 10.1074/jbc.M113.458315
Insights into β-lactamases from Burkholderia species, two phylogenetically related yet distinct resistance determinants
Abstract
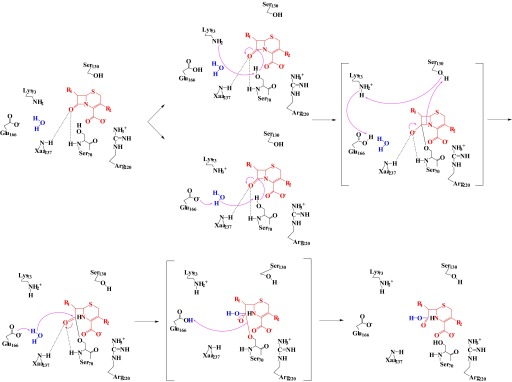
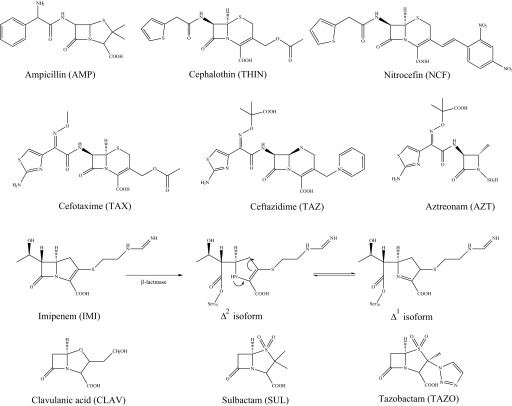
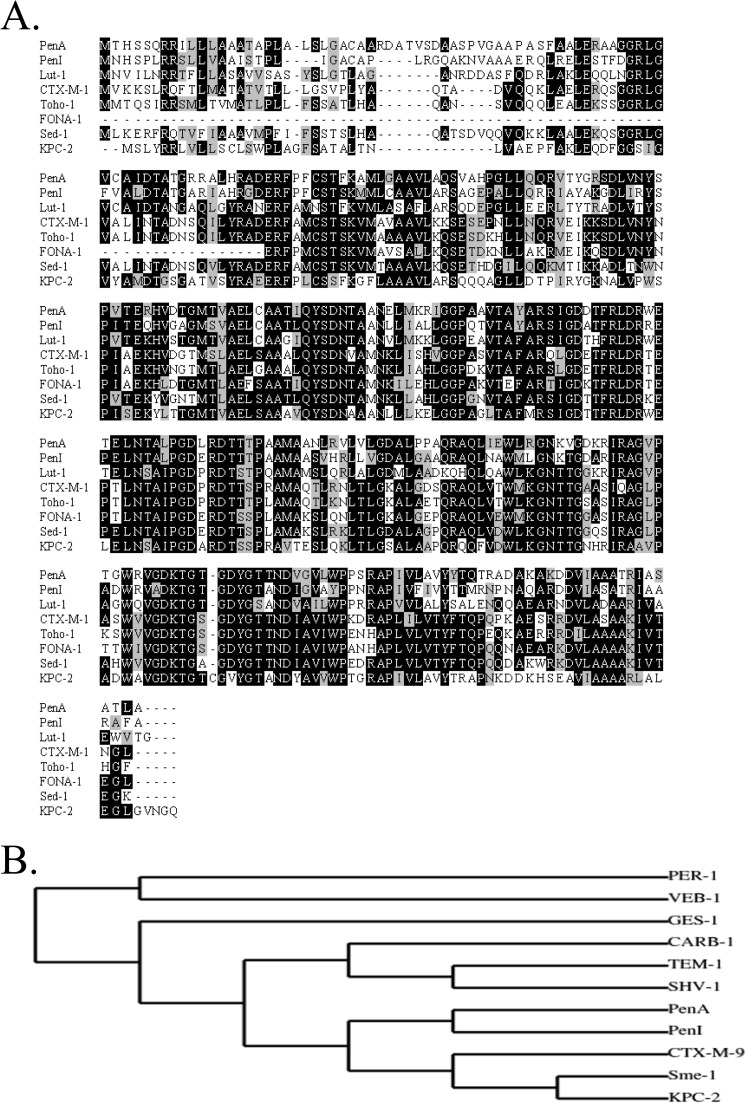
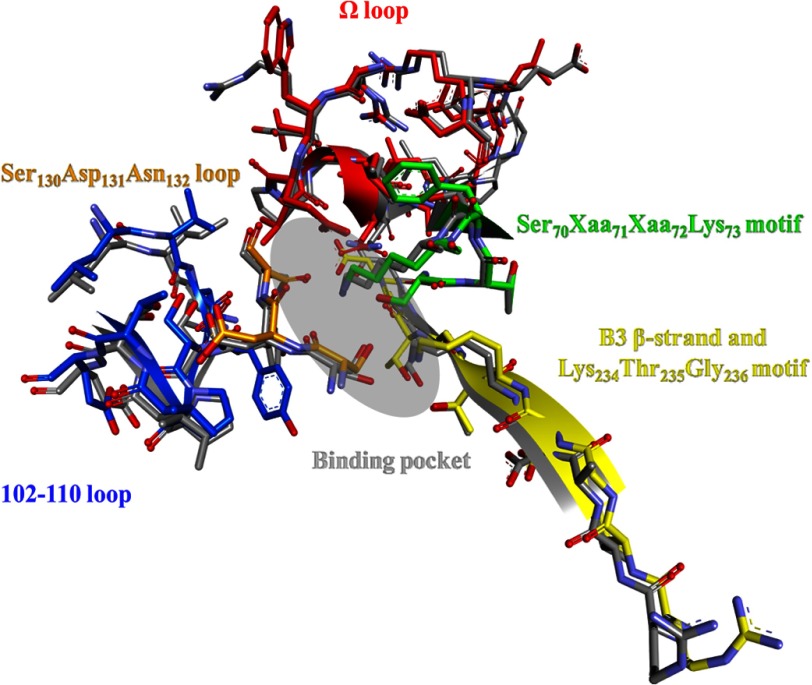
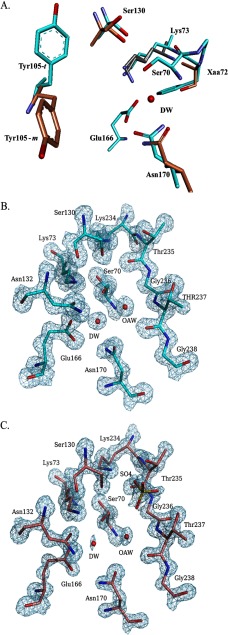
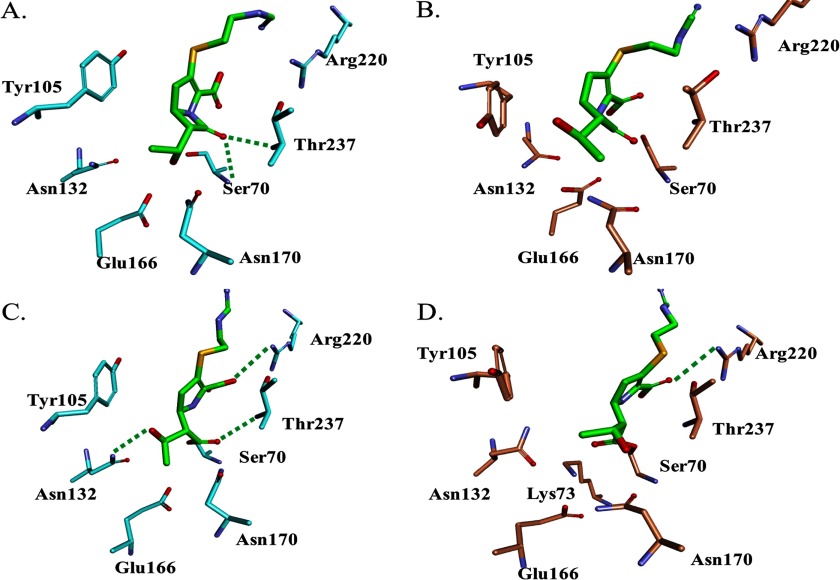
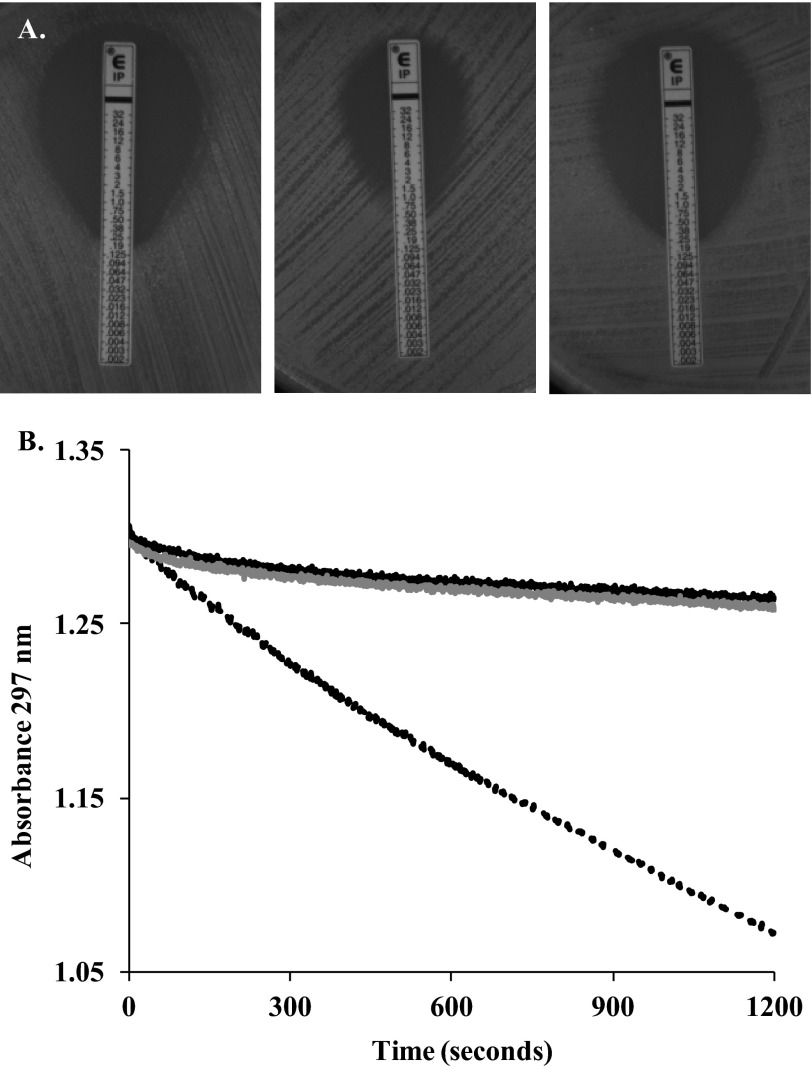
Burkholderia cepacia complex and Burkholderia pseudomallei are opportunistic human pathogens. Resistance to β-lactams among Burkholderia spp. is attributable to expression of β-lactamases (e.g. PenA in B. cepacia complex and PenI in B. pseudomallei). Phylogenetic comparisons reveal that PenA and PenI are highly related. However, the analyses presented here reveal that PenA is an inhibitor-resistant carbapenemase, most similar to KPC-2 (the most clinically significant serine carbapenemase), whereas PenI is an extended spectrum β-lactamase. PenA hydrolyzes β-lactams with k(cat) values ranging from 0.38 ± 0.04 to 460 ± 46 s(-1) and possesses high k(cat)/k(inact) values of 2000, 1500, and 75 for β-lactamase inhibitors. PenI demonstrates the highest kcat value for cefotaxime of 9.0 ± 0.9 s(-1). Crystal structure determination of PenA and PenI reveals important differences that aid in understanding their contrasting phenotypes. Changes in the positioning of conserved catalytic residues (e.g. Lys-73, Ser-130, and Tyr-105) as well as altered anchoring and decreased occupancy of the deacylation water explain the lower k(cat) values of PenI. The crystal structure of PenA with imipenem docked into the active site suggests why this carbapenem is hydrolyzed and the important role of Arg-220, which was functionally confirmed by mutagenesis and biochemical characterization. Conversely, the conformation of Tyr-105 hindered docking of imipenem into the active site of PenI. The structural and biochemical analyses of PenA and PenI provide key insights into the hydrolytic mechanisms of β-lactamases, which can lead to the rational design of novel agents against these pathogens.
Keywords: Antibiotic Resistance; Antibiotics; Microbiology; Molecular Modeling; Structural Biology.
Figures
References
-
- Mahenthiralingam E., Vandamme P. (2005) Taxonomy and pathogenesis of the Burkholderia cepacia complex. Chron. Respir. Dis. 2, 209–217 - PubMed
-
- Vandamme P., Dawyndt P. (2011) Classification and identification of the Burkholderia cepacia complex: past, present and future. Syst. Appl. Microbiol. 34, 87–95 - PubMed
-
- Mahenthiralingam E., Baldwin A., Dowson C. G. (2008) Burkholderia cepacia complex bacteria: opportunistic pathogens with important natural biology. J. Appl. Microbiol. 104, 1539–1551 - PubMed
-
- Suárez-Moreno Z. R., Caballero-Mellado J., Coutinho B. G., Mendonça-Previato L., James E. K., Venturi V. (2012) Common features of environmental and potentially beneficial plant-associated Burkholderia. Microb. Ecol. 63, 249–266 - PubMed
-
- Douce R. W., Zurita J., Sanchez O., Cardenas Aldaz P. (2008) Investigation of an outbreak of central venous catheter-associated bloodstream infection due to contaminated water. Infect. Control Hosp. Epidemiol. 29, 364–366 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
