

Polarized cell migration during cell-to-cell transmission of herpes simplex virus in human skin keratinocytes
- PMID: 23658449
- PMCID: PMC3700176
- DOI: 10.1128/JVI.01172-13
Polarized cell migration during cell-to-cell transmission of herpes simplex virus in human skin keratinocytes
Abstract
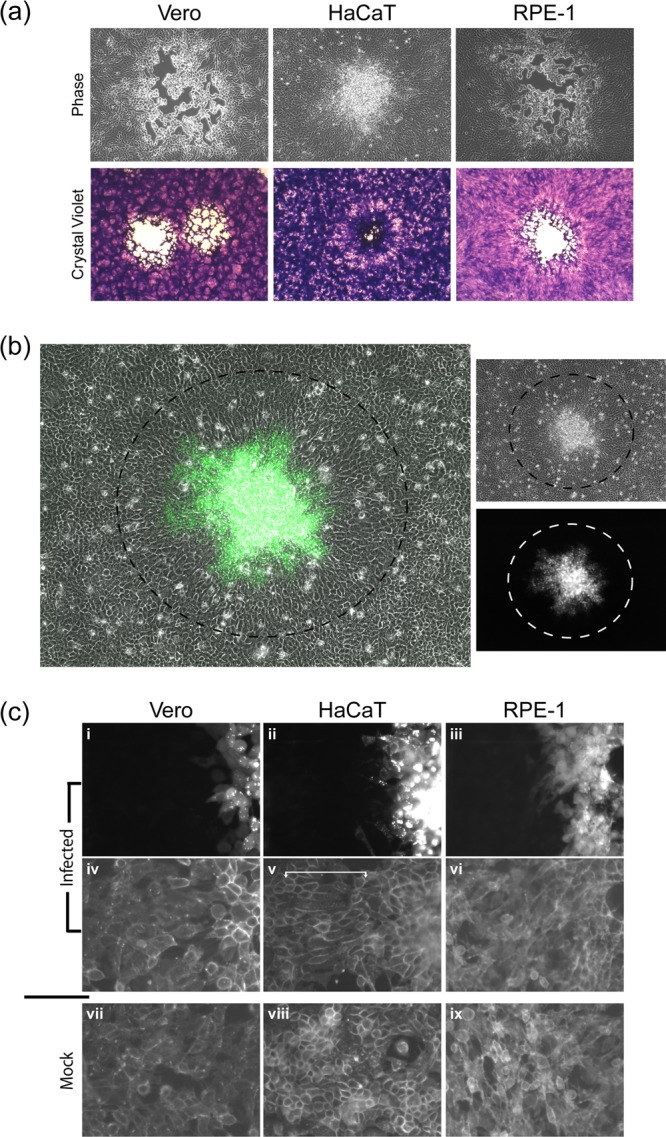
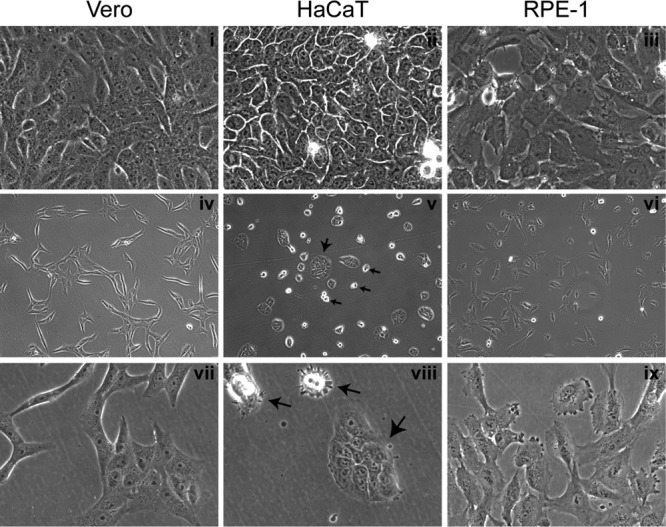
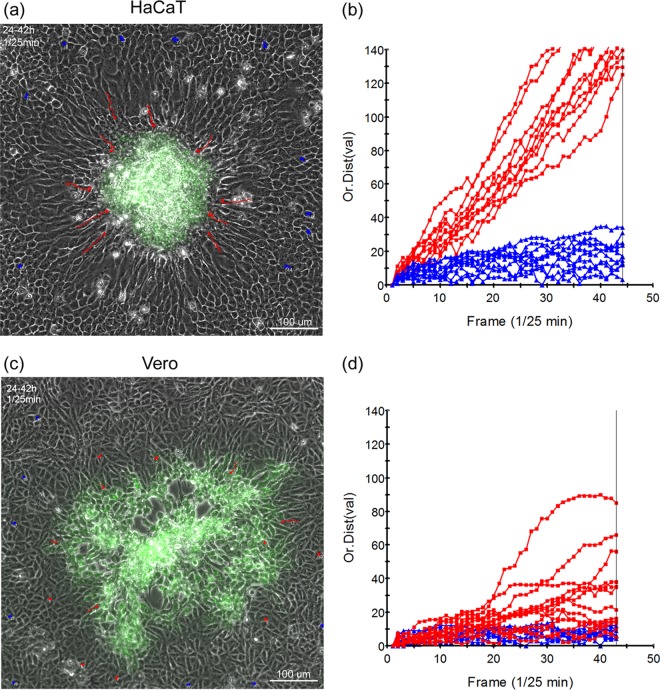
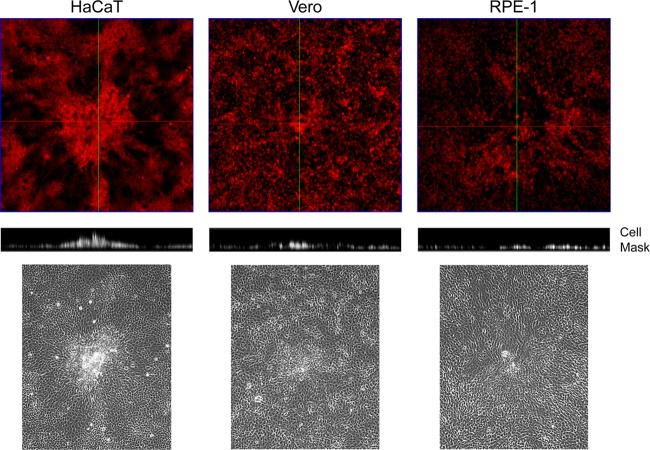
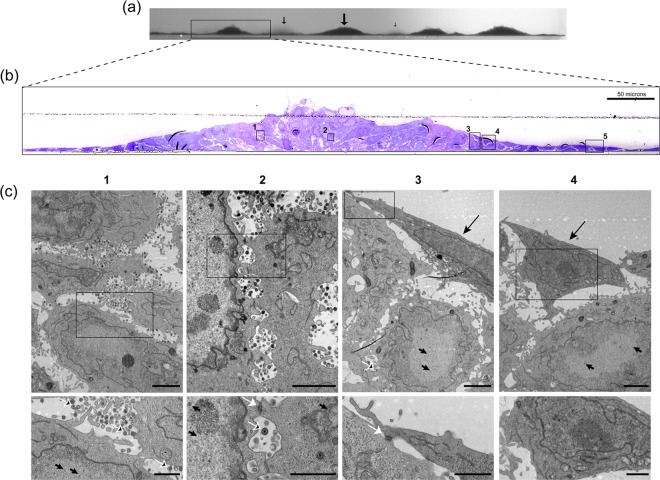
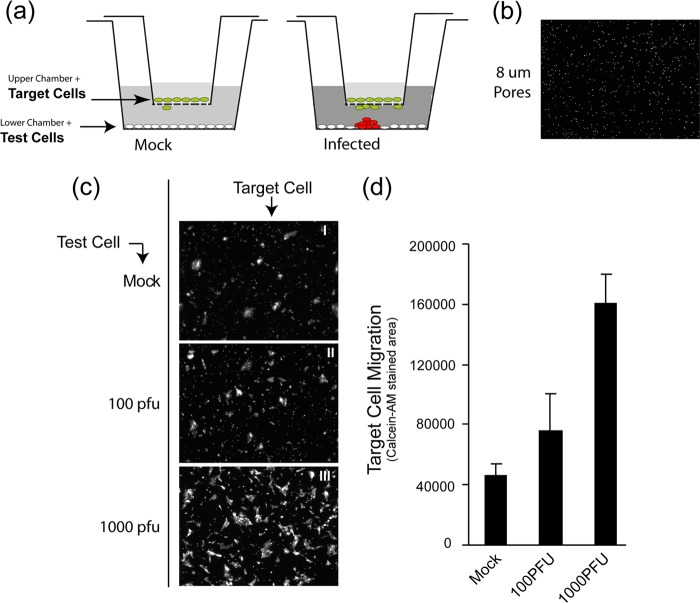
In addition to transmission involving extracellular free particles, a generally accepted model of virus propagation is one wherein virus replicates in one cell, producing infectious particles that transmit to the next cell via cell junctions or induced polarized contacts. This mechanism of spread is especially important in the presence of neutralizing antibody, and the concept underpins analysis of virus spread, plaque size, viral and host functions, and general mechanisms of virus propagation. Here, we demonstrate a novel process involved in cell-to-cell transmission of herpes simplex virus (HSV) in human skin cells that has not previously been appreciated. Using time-lapse microscopy of fluorescent viruses, we show that HSV infection induces the polarized migration of skin cells into the site of infection. In the presence of neutralizing antibody, uninfected skin cells migrate to the initial site of infection and spread over infected cells to become infected in a spatially confined cluster containing hundreds of cells. The cells in this cluster do not undergo cytocidal cell lysis but harbor abundant enveloped particles within cells and cell-free virus within interstitial regions below the cluster surface. Cells at the base and outside the cluster were generally negative for virus immediate-early expression. We further show, using spatially separated monolayer assays, that at least one component of this induced migration is the paracrine stimulation of a cytotactic response from infected cells to uninfected cells. The existence of this process changes our concept of virus transmission and the potential functions, virus, and host factors involved.
Figures


References
-
- Flint SJ, Enquist LW, Krug RM, Racaniello VR, Skalka AM. 2009. Principles of virology. ASM Press, Washington, DC
-
- Sattentau Q. 2008. Avoiding the void: cell-to-cell spread of human viruses. Nat. Rev. Microbiol. 6:815–826 - PubMed
-
- Black FL, Melnick JL. 1955. Microepidemiology of poliomyelitis and herpes-B infections: spread of the viruses within tissue cultures. J. Immunol. 74:236–242 - PubMed
-
- Christian RT, Ludovici PP, Jeter WS. 1971. Cell-to-cell transmission of herpes simplex virus in primary human amnion cells. Proc. Soc. Exp. Biol. Med. 138:1109–1115 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
