

Neuropeptides function in a homeostatic manner to modulate excitation-inhibition imbalance in C. elegans
- PMID: 23658528
- PMCID: PMC3642046
- DOI: 10.1371/journal.pgen.1003472
Neuropeptides function in a homeostatic manner to modulate excitation-inhibition imbalance in C. elegans
Abstract
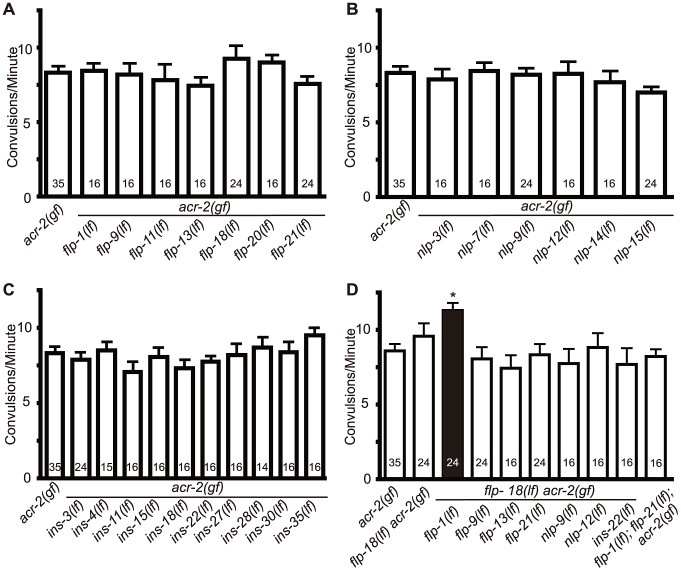
Neuropeptides play crucial roles in modulating neuronal networks, including changing intrinsic properties of neurons and synaptic efficacy. We previously reported a Caenorhabditis elegans mutant, acr-2(gf), that displays spontaneous convulsions as the result of a gain-of-function mutation in a neuronal nicotinic acetylcholine receptor subunit. The ACR-2 channel is expressed in the cholinergic motor neurons, and acr-2(gf) causes cholinergic overexcitation accompanied by reduced GABAergic inhibition in the locomotor circuit. Here we show that neuropeptides play a homeostatic role that compensates for this excitation-inhibition imbalance in the locomotor circuit. Loss of function in genes required for neuropeptide processing or release of dense core vesicles specifically modulate the convulsion frequency of acr-2(gf). The proprotein convertase EGL-3 is required in the cholinergic motor neurons to restrain convulsions. Electrophysiological recordings of neuromuscular junctions show that loss of egl-3 in acr-2(gf) causes a further reduction of GABAergic inhibition. We identify two neuropeptide encoding genes, flp-1 and flp-18, that together counteract the excitation-inhibition imbalance in acr-2(gf) mutants. We further find that acr-2(gf) causes an increased expression of flp-18 in the ventral cord cholinergic motor neurons and that overexpression of flp-18 reduces the convulsion of acr-2(gf) mutants. The effects of these peptides are in part mediated by two G-protein coupled receptors, NPR-1 and NPR-5. Our data suggest that the chronic overexcitation of the cholinergic motor neurons imposed by acr-2(gf) leads to an increased production of FMRFamide neuropeptides, which act to decrease the activity level of the locomotor circuit, thereby homeostatically modulating the excitation and inhibition imbalance.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






Similar articles
-
A neuronal acetylcholine receptor regulates the balance of muscle excitation and inhibition in Caenorhabditis elegans.PLoS Biol. 2009 Dec;7(12):e1000265. doi: 10.1371/journal.pbio.1000265. Epub 2009 Dec 22. PLoS Biol. 2009. PMID: 20027209 Free PMC article.
-
Neuronal transcriptome analyses reveal novel neuropeptide modulators of excitation and inhibition imbalance in C. elegans.PLoS One. 2020 Jun 4;15(6):e0233991. doi: 10.1371/journal.pone.0233991. eCollection 2020. PLoS One. 2020. PMID: 32497060 Free PMC article.
-
Loss of neuropeptidergic regulation of cholinergic transmission induces homeostatic compensation in muscle cells to preserve synaptic strength.PLoS Biol. 2025 May 8;23(5):e3003171. doi: 10.1371/journal.pbio.3003171. eCollection 2025 May. PLoS Biol. 2025. PMID: 40338987 Free PMC article.
-
Neuropeptide gene families in the nematode Caenorhabditis elegans.Ann N Y Acad Sci. 1999;897:239-52. doi: 10.1111/j.1749-6632.1999.tb07895.x. Ann N Y Acad Sci. 1999. PMID: 10676452 Review.
-
Neuropeptides.WormBook. 2008 Sep 25:1-36. doi: 10.1895/wormbook.1.142.1. WormBook. 2008. PMID: 18819171 Free PMC article. Review.
Cited by
-
Homeostasis in C. elegans sleep is characterized by two behaviorally and genetically distinct mechanisms.Elife. 2014 Dec 4;3:e04380. doi: 10.7554/eLife.04380. Elife. 2014. PMID: 25474127 Free PMC article.
-
A molecular atlas of adult C. elegans motor neurons reveals ancient diversity delineated by conserved transcription factor codes.Cell Rep. 2024 Mar 26;43(3):113857. doi: 10.1016/j.celrep.2024.113857. Epub 2024 Feb 29. Cell Rep. 2024. PMID: 38421866 Free PMC article.
-
Annotation of nerve cord transcriptome in earthworm Eisenia fetida.Genom Data. 2017 Oct 12;14:91-105. doi: 10.1016/j.gdata.2017.10.002. eCollection 2017 Dec. Genom Data. 2017. PMID: 29204349 Free PMC article.
-
Neuroendocrine Control of Synaptic Transmission by PHAC-1 in C. elegans.J Neurosci. 2025 Mar 26;45(13):e1767232024. doi: 10.1523/JNEUROSCI.1767-23.2024. J Neurosci. 2025. PMID: 39919830
-
Regulation of Glutamate Signaling in the Sensorimotor Circuit by CASY-1A/Calsyntenin in Caenorhabditis elegans.Genetics. 2018 Apr;208(4):1553-1564. doi: 10.1534/genetics.118.300834. Epub 2018 Feb 23. Genetics. 2018. PMID: 29475851 Free PMC article.
References
-
- Krieger DT (1983) Brain peptides: what, where, and why? Science 222: 975–985. - PubMed
-
- Kow LM, Pfaff DW (1988) Neuromodulatory actions of peptides. Annu Rev Pharmacol Toxicol 28: 163–188. - PubMed
-
- Blake AD, Badway AC, Strowski MZ (2004) Delineating somatostatin's neuronal actions. Curr Drug Targets CNS Neurol Disord 3: 153–160. - PubMed
-
- Fetissov SO, Jacoby AS, Brumovsky PR, Shine J, Iismaa TP, et al. (2003) Altered hippocampal expression of neuropeptides in seizure-prone GALR1 knockout mice. Epilepsia 44: 1022–1033. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
