

Downregulation of OPA3 is responsible for transforming growth factor-β-induced mitochondrial elongation and F-actin rearrangement in retinal pigment epithelial ARPE-19 cells
- PMID: 23658835
- PMCID: PMC3643898
- DOI: 10.1371/journal.pone.0063495
Downregulation of OPA3 is responsible for transforming growth factor-β-induced mitochondrial elongation and F-actin rearrangement in retinal pigment epithelial ARPE-19 cells
Abstract
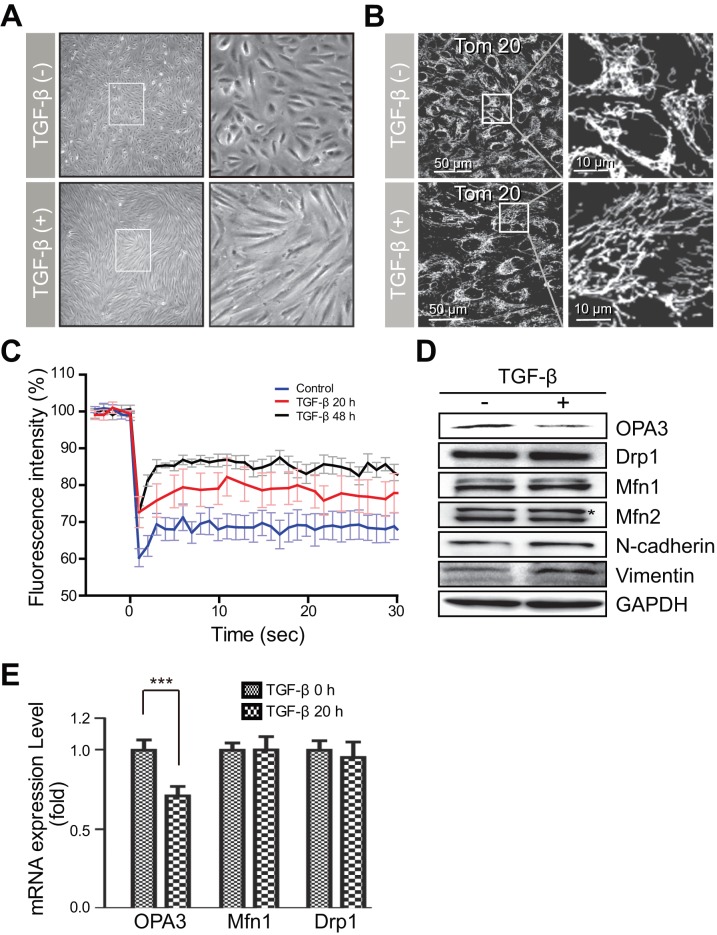
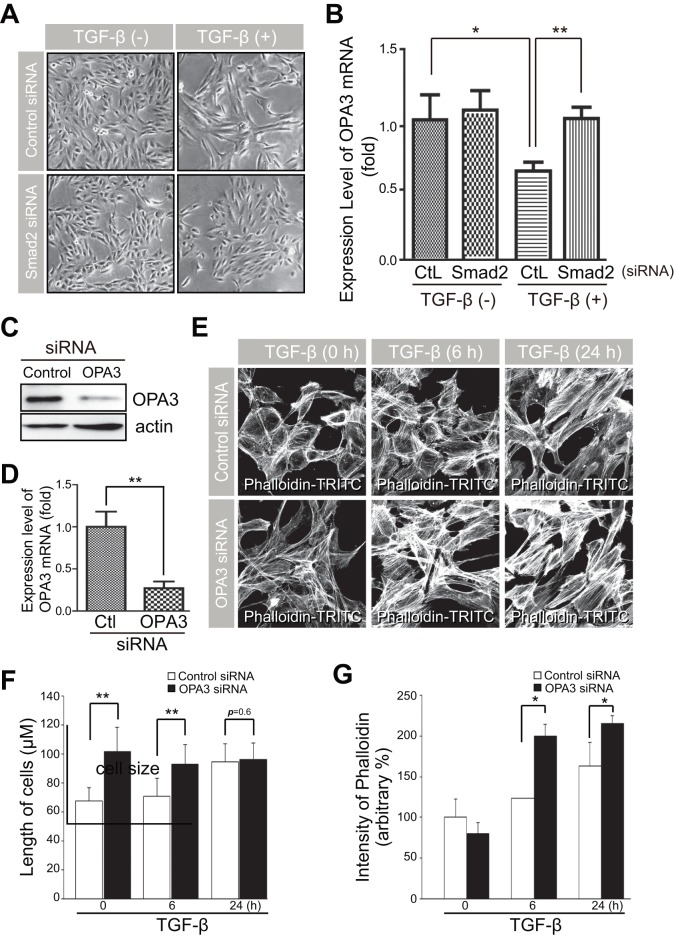
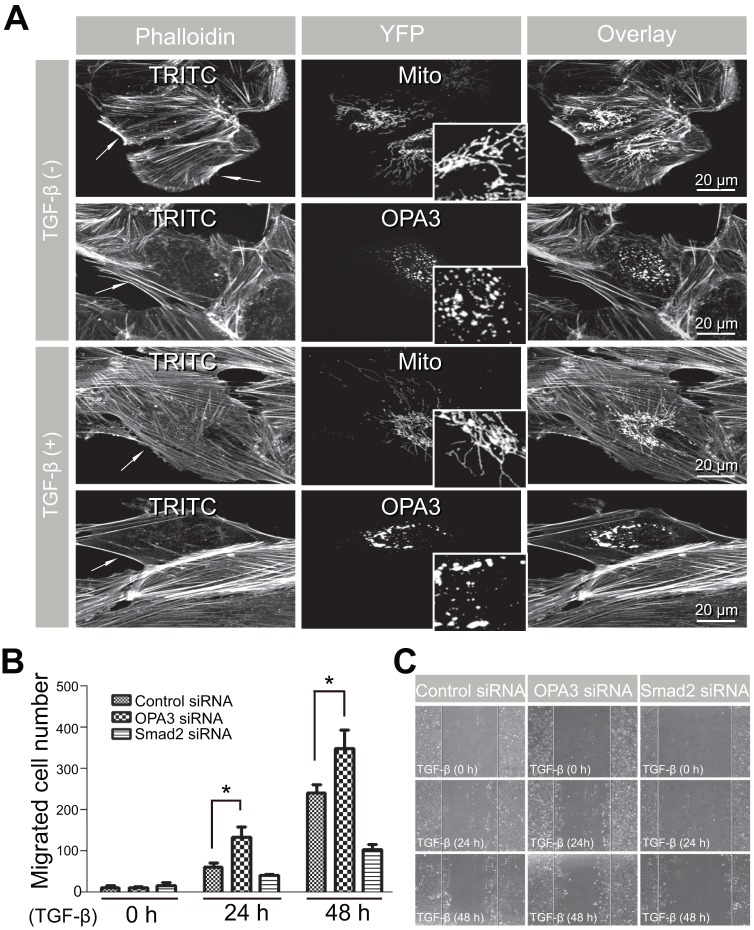
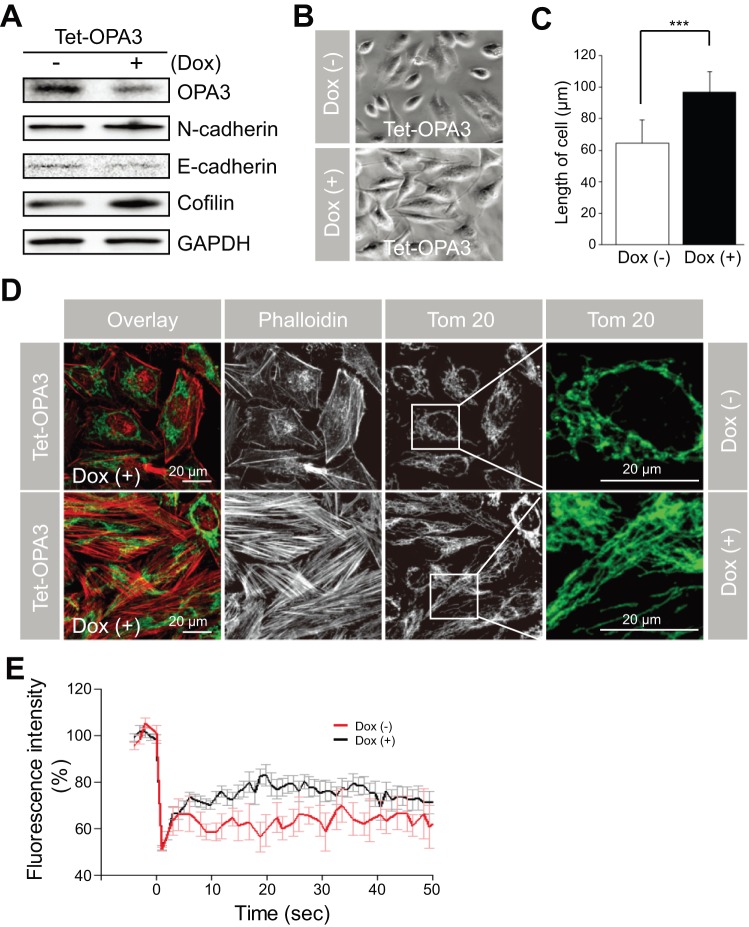
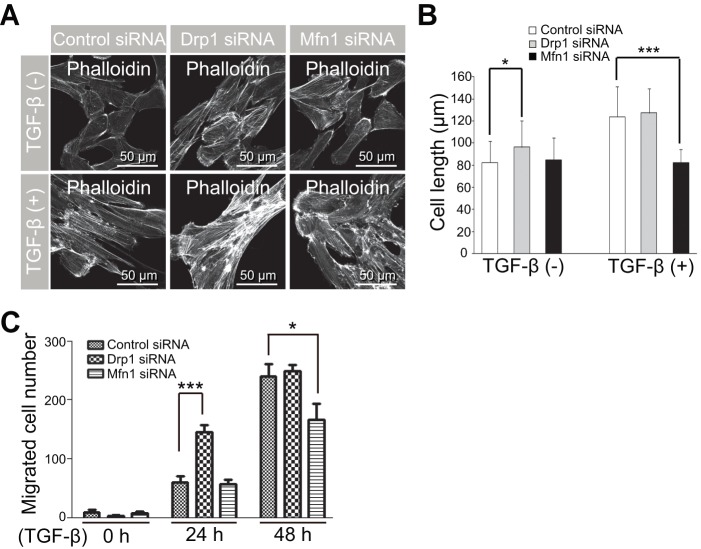
Transforming growth factor-β signaling is known to be a key signaling pathway in the induction of epithelial-mesenchymal transition. However, the mechanism of TGF-β signaling in the modulation of EMT remains unclear. In this study, we found that TGF-β treatment resulted in elongation of mitochondria accompanied by induction of N-cadherin, vimentin, and F-actin in retinal pigment epithelial cells. Moreover, OPA3, which plays a crucial role in mitochondrial dynamics, was downregulated following TGF-β treatment. Suppression of TGF-β signaling using Smad2 siRNA prevented loss of OPA3 induced by TGF-β. Knockdown of OPA3 by siRNA and inducible shRNA significantly increased stress fiber levels, cell length, cell migration and mitochondrial elongation. In contrast, forced expression of OPA3 in ARPE-19 cells inhibited F-actin rearrangement and induced mitochondrial fragmentation. We also showed that Drp1 depletion increased cell length and induced rearrangement of F-actin. Depletion of Mfn1 blocked the increase in cell length during TGF-β-mediated EMT. These results collectively substantiate the involvement of mitochondrial dynamics in TGF-β-induced EMT.
Conflict of interest statement
Figures
References
-
- Feng XH, Derynck R (2005) Specificity and versatility in tgf-beta signaling through Smads. Annu Rev Cell Dev Biol 21: 659–693. - PubMed
-
- Massague J, Seoane J, Wotton D (2005) Smad transcription factors. Genes Dev 19: 2783–2810. - PubMed
-
- Groppe J, Hinck CS, Samavarchi-Tehrani P, Zubieta C, Schuermann JP, et al. (2008) Cooperative assembly of TGF-beta superfamily signaling complexes is mediated by two disparate mechanisms and distinct modes of receptor binding. Mol Cell 29: 157–168. - PubMed
-
- Thiery JP (2003) Epithelial-mesenchymal transitions in development and pathologies. Curr Opin Cell Biol 15: 740–746. - PubMed
-
- Heldin CH, Landstrom M, Moustakas A (2009) Mechanism of TGF-beta signaling to growth arrest, apoptosis, and epithelial-mesenchymal transition. Curr Opin Cell Biol 21: 166–176. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
