

Differential roles of human Dicer-binding proteins TRBP and PACT in small RNA processing
- PMID: 23661684
- PMCID: PMC3711433
- DOI: 10.1093/nar/gkt361
Differential roles of human Dicer-binding proteins TRBP and PACT in small RNA processing
Abstract
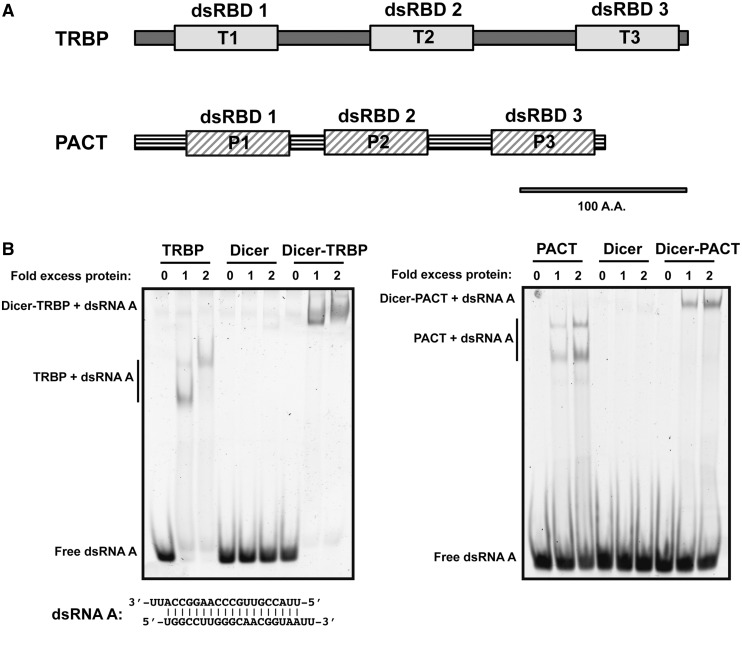
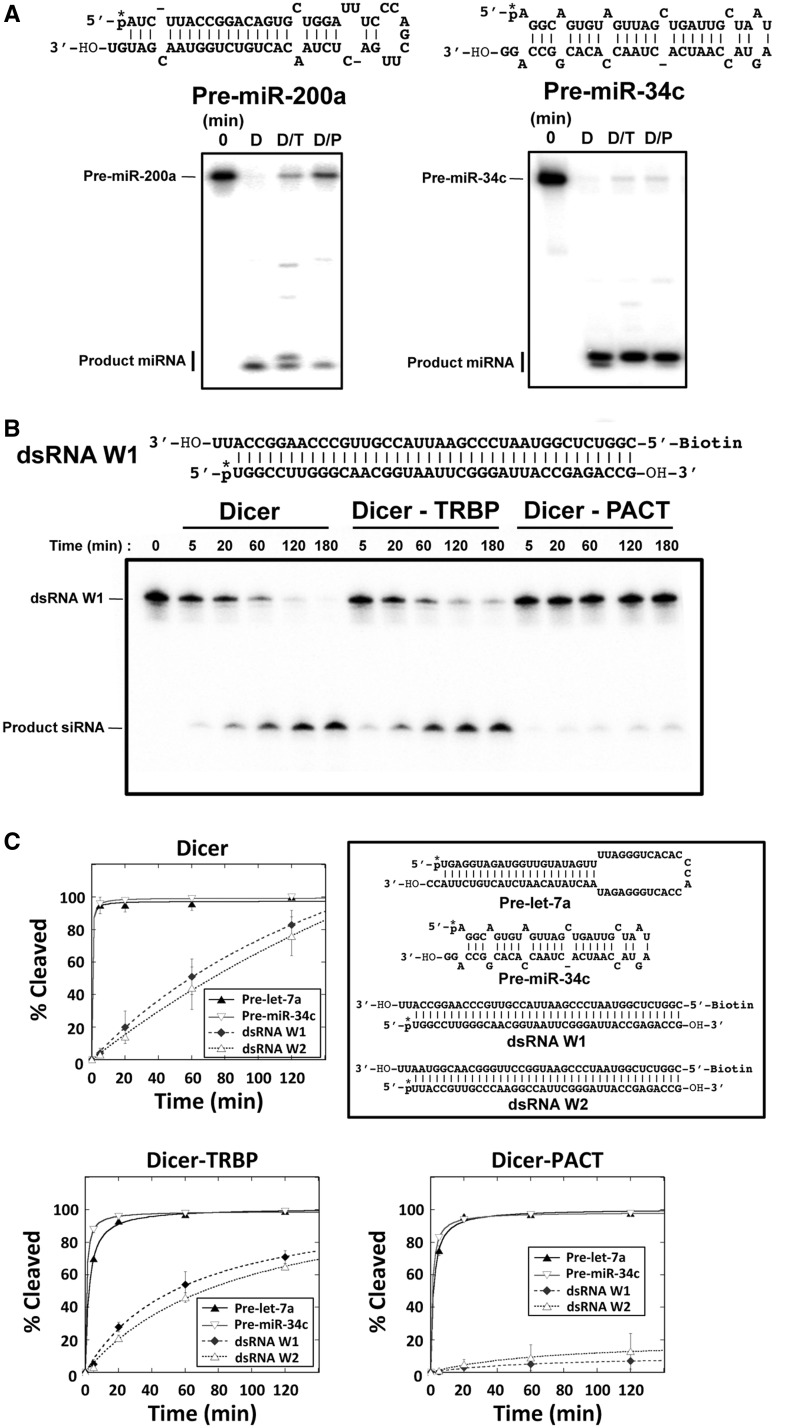
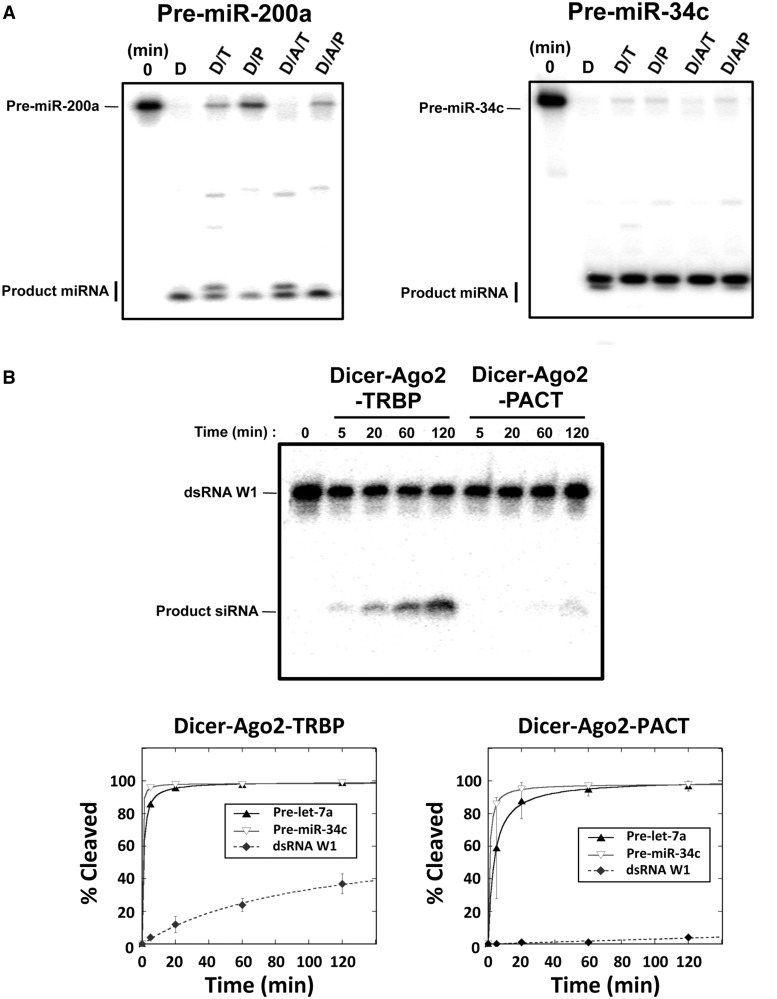
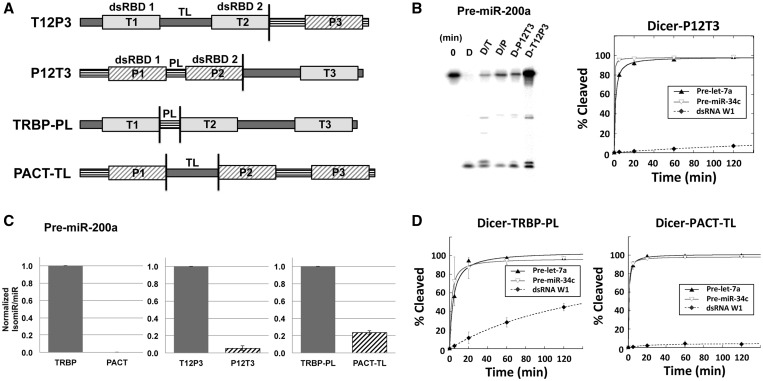
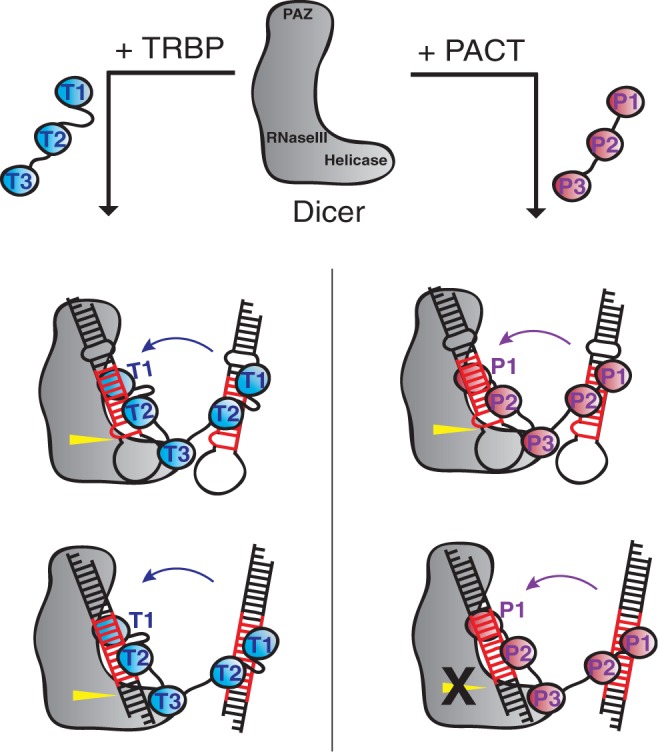
During RNA interference and related gene regulatory pathways, the endonuclease Dicer cleaves precursor RNA molecules to produce microRNAs (miRNAs) and short interfering RNAs (siRNAs). Human cells encode a single Dicer enzyme that can associate with two different double-stranded RNA (dsRNA)-binding proteins, protein activator of PKR (PACT) and trans-activation response RNA-binding protein (TRBP). However, the functional redundancy or differentiation of PACT and TRBP in miRNA and siRNA biogenesis is not well understood. Using a reconstituted system, we show here that PACT and TRBP have distinct effects on Dicer-mediated dsRNA processing. In particular, we found that PACT in complex with Dicer inhibits the processing of pre-siRNA substrates when compared with Dicer and a Dicer-TRBP complex. In addition, PACT and TRBP show non-redundant effects on the production of different-sized miRNAs (isomiRs), which in turn alter target-binding specificities. Experiments using chimeric versions of PACT and TRBP suggest that the two N-terminal RNA-binding domains of each protein confer the observed differences in dsRNA substrate recognition and processing behavior of Dicer-dsRNA-binding protein complexes. These results support the conclusion that in humans, Dicer-associated dsRNA-binding proteins are important regulatory factors that contribute both substrate and cleavage specificity during miRNA and siRNA production.
Figures
References
-
- Kim VN, Han J, Siomi MC. Biogenesis of small RNAs in animals. Nat. Rev. Mol. Cell. Biol. 2009;10:126–139. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
