

doi: 10.1016/j.neuron.2013.02.030.
The cell-autonomous role of excitatory synaptic transmission in the regulation of neuronal structure and function
Affiliations
- PMID: 23664612
- PMCID: PMC3666354
- DOI: 10.1016/j.neuron.2013.02.030
Item in Clipboard
The cell-autonomous role of excitatory synaptic transmission in the regulation of neuronal structure and function
Neuron.
.
Abstract
The cell-autonomous role of synaptic transmission in the regulation of neuronal structural and electrical properties is unclear. We have now employed a genetic approach to eliminate glutamatergic synaptic transmission onto individual CA1 pyramidal neurons in a mosaic fashion in vivo. Surprisingly, while electrical properties are profoundly affected in these neurons, as well as inhibitory synaptic transmission, we found little perturbation of neuronal morphology, demonstrating a functional segregation of excitatory synaptic transmission from neuronal morphological development.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
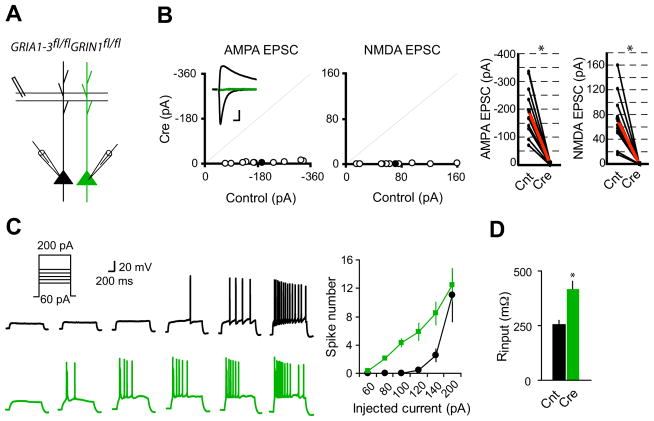
(A) Schematic of dual whole-cell voltage-clamp recording in acute hippocampal slices from GRIA1-3fl/flGRIN1fl/fl p20-p28 mice infected with virus expressing CreGFP at p0. (B) Scatter plots showed amplitudes of EPSCs for single pairs (open circles) and mean ± SEM (filled circles). (Inset) Representative superimposed traces (Black, Control (Cnt); Green, Cre). Scale bar: 0.02 s, 50 pA. Graphs in the right show that synaptic transmission mediated by AMPARs or NMDARs is essentially eliminated in CreGFP-expressing neurons (each black line represents a single pair and the red line represents the average; AMPA EPSCs, Cnt: −194.2 ± 29.8 pA; Cre: −4.92 ± 1.39 pA; n = 10; NMDA EPSCS, Cnt: 71.5 ± 14.03 pA; Cre: 0.56 ± 0.08 pA; n = 10). *P < 0.001. (C) Sample action potential responses to step current injections (60, 80, 100, 120, 140 and 200 pA; 1000 ms) in control neurons (black) or CreGFP-expressing neurons (Green). Right is the summary graph showing that deletion of both AMPARs and NMDARs increased the excitability of CA1 pyramidal neurons. *P < 0.001. (D) Bar graph shows that input resistance is significantly enhanced in neurons lacking both AMPARs and NMDARs. *P < 0.001.
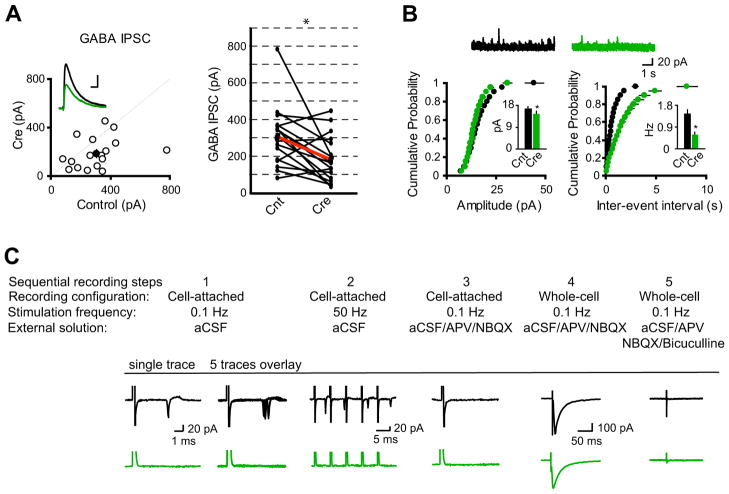
(A) Scatter plots showed amplitudes of IPSCs for single pairs (open circles) and mean ± SEM (filled circles). (Inset) Representative superimposed traces (Black, Cnt; Green, Cre). Scale bar: 0.05 s, 100 pA. Graph in the right shows that synaptic transmission mediated by GABAA receptors is significantly reduced in CreGFP-expressing neurons (each black line represents a single pair and the red line represents the average; Cnt: 309.00 ± 39.32 pA; Cre: 183.53 ± 32.13 pA; n = 17). *P < 0.001. (B) Cumulative distributions of mIPSC amplitudes and inter-event intervals from control and CreGFP-expressing neurons. Above are sample traces from a control neuron (Black) and a CreGFP-expressing neuron (Green). Bar graphs show that both mIPSC amplitudes and inter-event intervals are reduced in neurons expressing CreGFP. *P < 0.05. (C) Synaptic activation-evoked action potential firing is lost in CA1 pyramidal neurons expressing CreGFP. Synaptic activation-evoked action potentials were simultaneously recorded in one CreGFP expressing neuron and one neighboring control neuron under cell-attached mode without breaking-in. Synaptic activity was induced by stimulation of Schaffer Collateral pathway at either 0.1 Hz (step 1) or 50 Hz (step 2). After washing-in AP-V (100 μM) and NBQX (10 μM), synaptic activation-evoked action potentials were blocked (step 3). Neurons were then held in whole-cell mode under voltage-clamp configuration to record evoked IPSCs (step 4), which were completely blocked by 20 μM bicuculline (step 5).
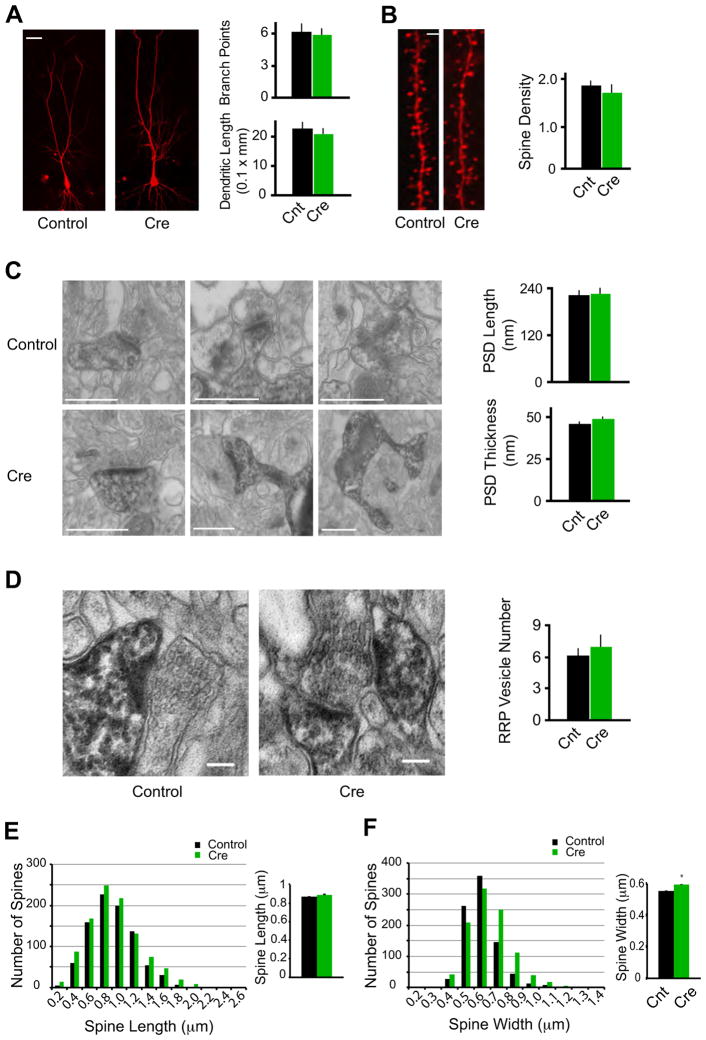
Morphological conservation of CA1 pyramidal neurons devoid of synaptic excitation in vivo. (A) Representative confocal stacks from Cnt and Cre cells. Bar graph in right shows average number dendritic branch points and dendritic length (Cnt: n = 10; Cre: n = 8; P > 0.05). Scale bar, 20 μm. (B) Maximum intensity projections of representative dendrites from Cnt and Cre cells. Bar graph in right shows average spine density (the number of spines per micron) for Cnt and Cre neurons (Cnt: n = 7 neurons; Cre: n = 6; P > 0.05). Scale bar, 1 μm. (C) Electron micrographs consistently reveal PSDs in spines from Cre-expressing cell, similar to Cnt cells. Bar graphs show no significant difference in PSD length or thickness between Cnt and Cre spines (Cnt: n = 33 spines; Cre: n = 40; P > 0.05). Scale bars, 500 nm. (D) Pre-synaptic vesicle pools in boutons opposed to Cnt and Cre spines. Bar graph shows no significant difference in RRP vesicle numbers (Cnt: n = 15 spines; Cre: n = 15; P > 0.05). Scale bars, 100 nm. (E–F) Histograms showing spine lengths (E) and widths (F) for all spines measured in confocal volumes of control (Black) and Cre-expressing (Green) neurons. In the bar graphs in the right, comparing the two conditions revealed no significant difference in mean spine length (E, Cnt: 0.87 ± 0.01 μm, n = 880; Cre: 0.88 ± 0.01 μm, n = 1021; P = 0.23), and a small but statistically significant increase in spine width in Cre-expressing neurons (F, Cnt: 0.55 ± 0.004 μm, n = 861; Cre: 0.59 ± 0.004 μm, n = 998; P < 0.01).
References
-
- Ben-Ari Y. Excitatory actions of gaba during development: the nature of the nurture. Nature reviews Neuroscience. 2002;3:728–739. - PubMed
-
- Ben-Ari Y, Gaiarsa JL, Tyzio R, Khazipov R. GABA: a pioneer transmitter that excites immature neurons and generates primitive oscillations. Physiological reviews. 2007;87:1215–1284. - PubMed
-
- Bhatt DH, Zhang S, Gan WB. Dendritic spine dynamics. Annu Rev Physiol. 2009;71:261–282. - PubMed
-
- Bindu B, Alladi PA, Mansooralikhan BM, Srikumar BN, Raju TR, Kutty BM. Short-term exposure to an enriched environment enhances dendritic branching but not brain-derived neurotrophic factor expression in the hippocampus of rats with ventral subicular lesions. Neuroscience. 2007;144:412–423. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
