

Macropinocytosis of protein is an amino acid supply route in Ras-transformed cells
- PMID: 23665962
- PMCID: PMC3810415
- DOI: 10.1038/nature12138
Macropinocytosis of protein is an amino acid supply route in Ras-transformed cells
Abstract
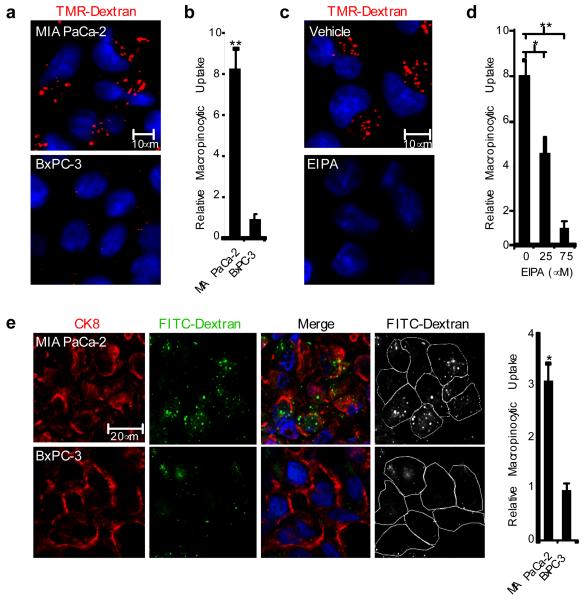
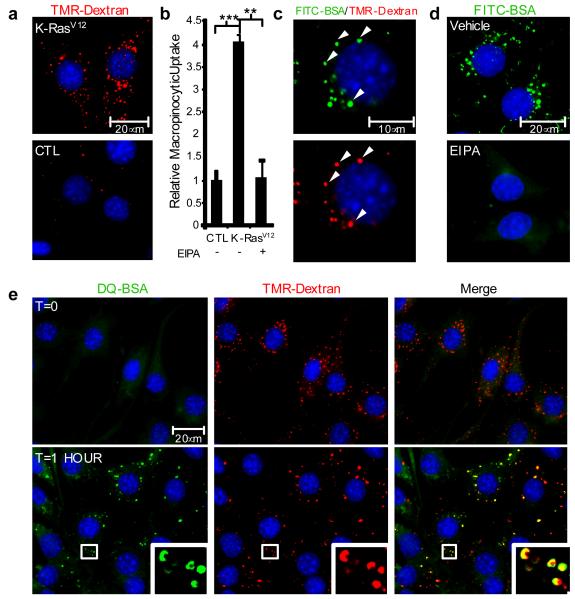
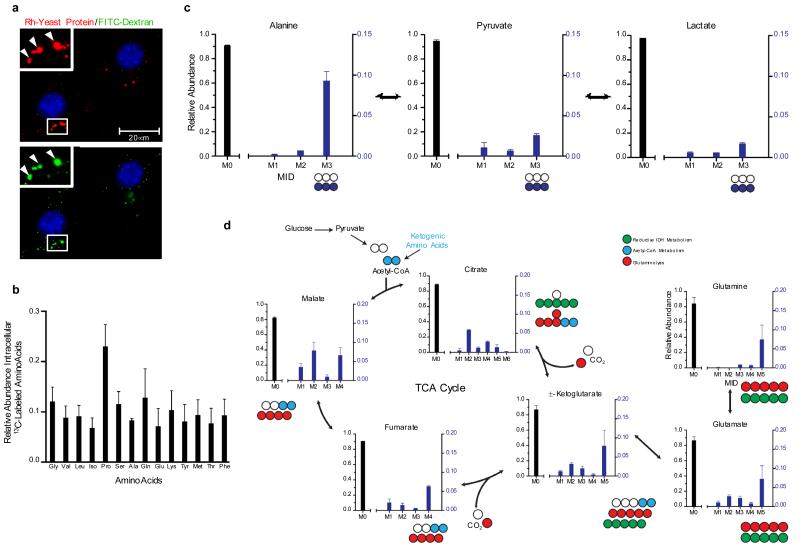
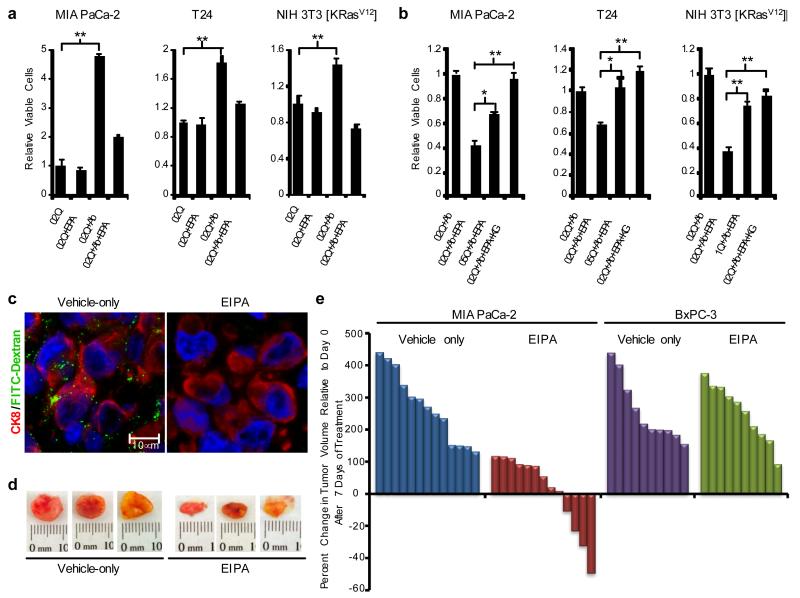
Macropinocytosis is a highly conserved endocytic process by which extracellular fluid and its contents are internalized into cells through large, heterogeneous vesicles known as macropinosomes. Oncogenic Ras proteins have been shown to stimulate macropinocytosis but the functional contribution of this uptake mechanism to the transformed phenotype remains unknown. Here we show that Ras-transformed cells use macropinocytosis to transport extracellular protein into the cell. The internalized protein undergoes proteolytic degradation, yielding amino acids including glutamine that can enter central carbon metabolism. Accordingly, the dependence of Ras-transformed cells on free extracellular glutamine for growth can be suppressed by the macropinocytic uptake of protein. Consistent with macropinocytosis representing an important route of nutrient uptake in tumours, its pharmacological inhibition compromises the growth of Ras-transformed pancreatic tumour xenografts. These results identify macropinocytosis as a mechanism by which cancer cells support their unique metabolic needs and point to the possible exploitation of this process in the design of anticancer therapies.
Figures
Comment in
-
Metabolism: Taking it all in.Nat Rev Cancer. 2013 Jul;13(7):438. doi: 10.1038/nrc3550. Epub 2013 Jun 6. Nat Rev Cancer. 2013. PMID: 23739049 No abstract available.
References
-
- Bar-Sagi D, Feramisco JR. Induction of membrane ruffling and fluid-phase pinocytosis in quiescent fibroblasts by ras proteins. Science. 1986;233:1061–1068. - PubMed
-
- Walsh AB, Bar-Sagi D. Differential activation of the Rac pathway by Ha-Ras and K-Ras. J Biol Chem. 2001;276:15609–15615. - PubMed
-
- Lopez-Crapez E, Chypre C, Saavedra J, Marchand J, Grenier J. Rapid and large-scale method to detect K-ras gene mutations in tumor samples. Clinical chemistry. 1997;43:936–942. - PubMed
-
- Aoki K, Yoshida T, Sugimura T, Terada M. Liposome-mediated in vivo gene transfer of antisense K-ras construct inhibits pancreatic tumor dissemination in the murine peritoneal cavity. Cancer Res. 1995;55:3810–3816. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
