

Multiplexing stimulus information through rate and temporal codes in primate somatosensory cortex
- PMID: 23667327
- PMCID: PMC3646728
- DOI: 10.1371/journal.pbio.1001558
Multiplexing stimulus information through rate and temporal codes in primate somatosensory cortex
Abstract
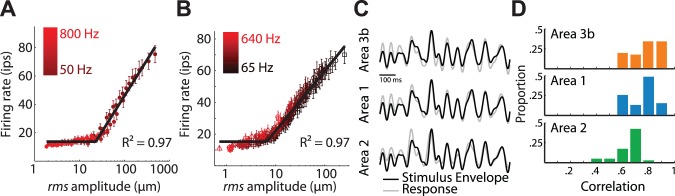
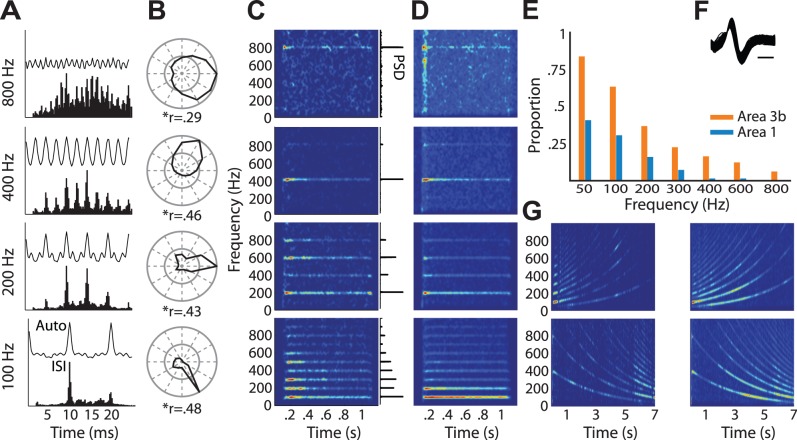
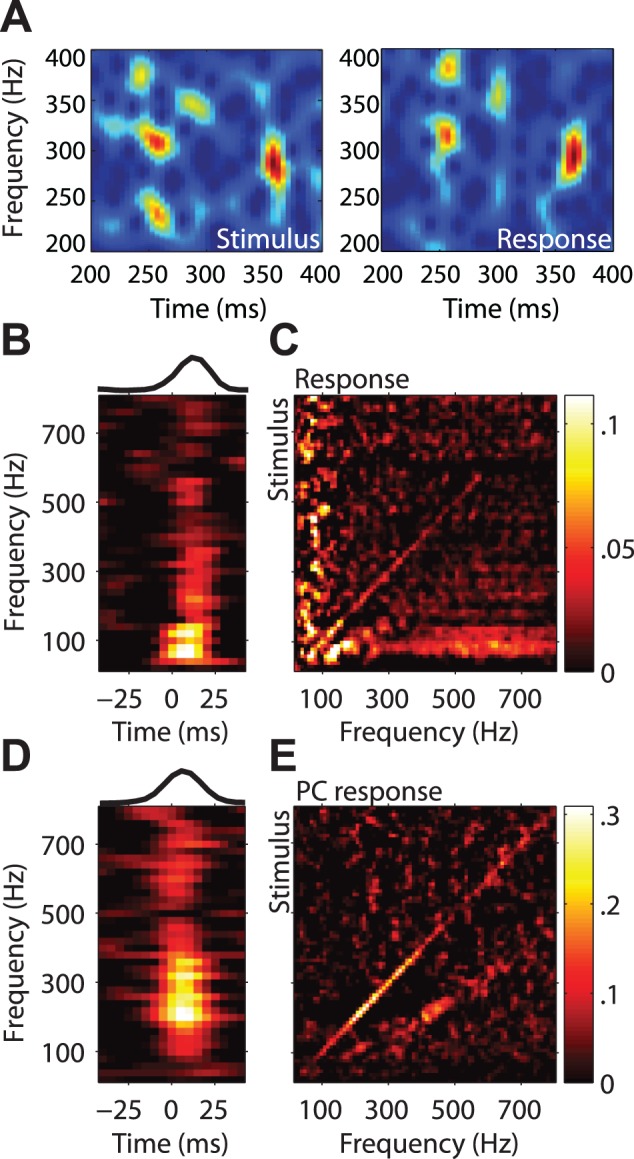
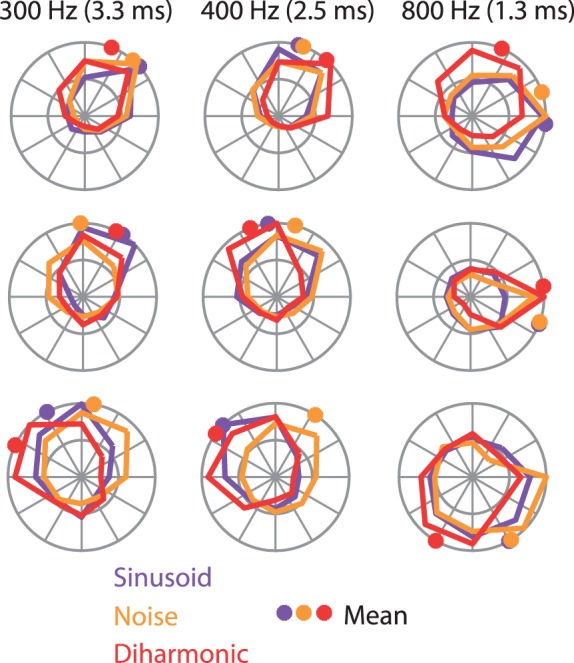
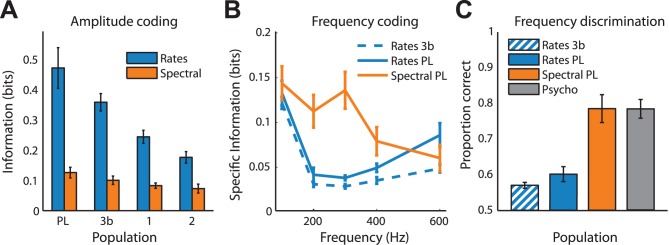
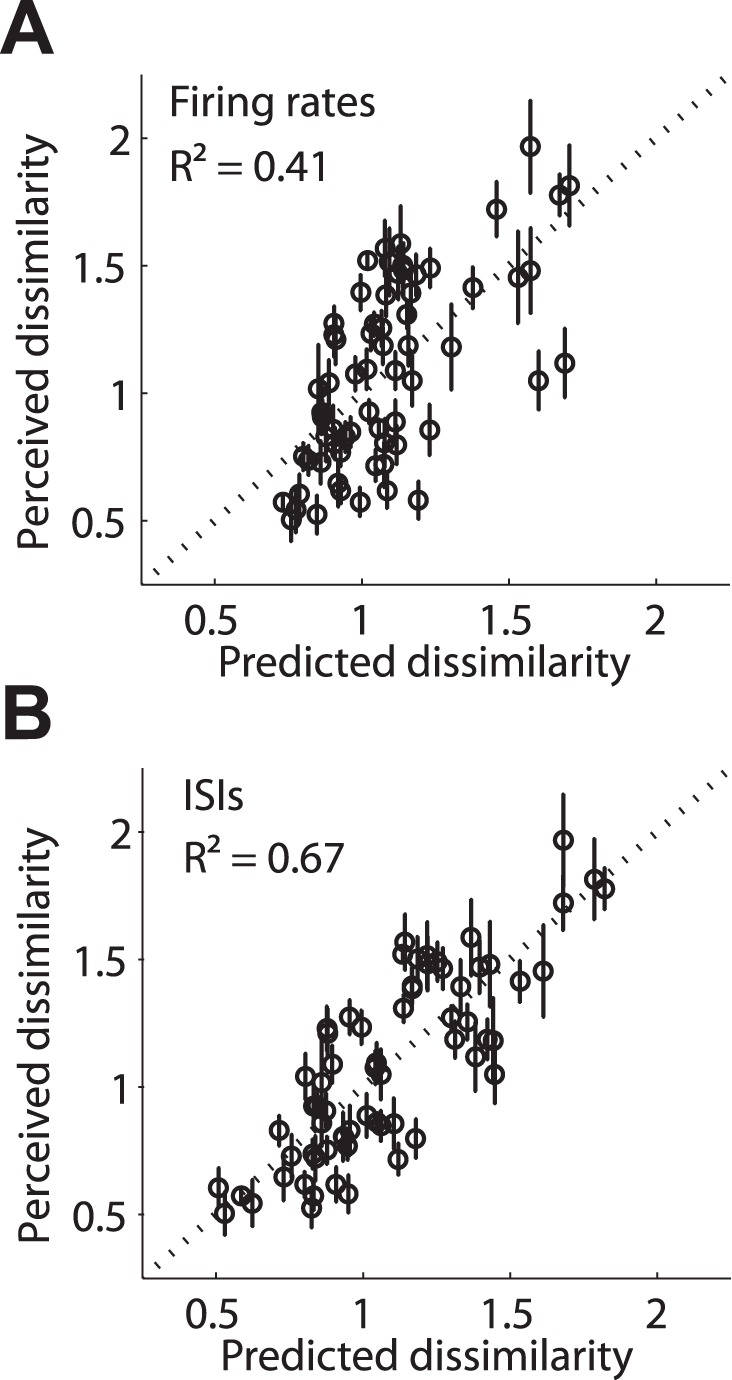
Our ability to perceive and discriminate textures relies on the transduction and processing of complex, high-frequency vibrations elicited in the fingertip as it is scanned across a surface. How naturalistic vibrations, and by extension texture, are encoded in the responses of neurons in primary somatosensory cortex (S1) is unknown. Combining single unit recordings in awake macaques and perceptual judgments obtained from human subjects, we show that vibratory amplitude is encoded in the strength of the response evoked in S1 neurons. In contrast, the frequency composition of the vibrations, up to 800 Hz, is not encoded in neuronal firing rates, but rather in the phase-locked responses of a subpopulation of neurons. Moreover, analysis of perceptual judgments suggests that spike timing not only conveys stimulus information but also shapes tactile perception. We conclude that information about the amplitude and frequency of natural vibrations is multiplexed at different time scales in S1, and encoded in the rate and temporal patterning of the response, respectively.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Hollins M, Risner SR (2000) Evidence for the duplex theory of tactile texture perception. Percept Psychophys 62: 695–705. - PubMed
-
- Bensmaia S, Hollins M (2005) Pacinian representations of fine surface texture. Percept Psychophys 67: 842–854. - PubMed
-
- Weber AI, Cheng JW, Dammann JF, Bensmaia SJ (2011) The coding of natural textures at the somatosensory periphery. In: 41st Annual Meeting of the Society for Neuroscience, Washington (D.C.), Abstract 750.01.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
