

Essential role for Cdk2 inhibitory phosphorylation during replication stress revealed by a human Cdk2 knockin mutation
- PMID: 23671119
- PMCID: PMC3670391
- DOI: 10.1073/pnas.1302927110
Essential role for Cdk2 inhibitory phosphorylation during replication stress revealed by a human Cdk2 knockin mutation
Abstract
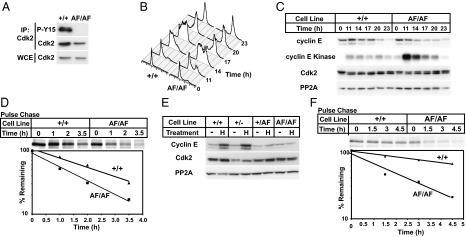
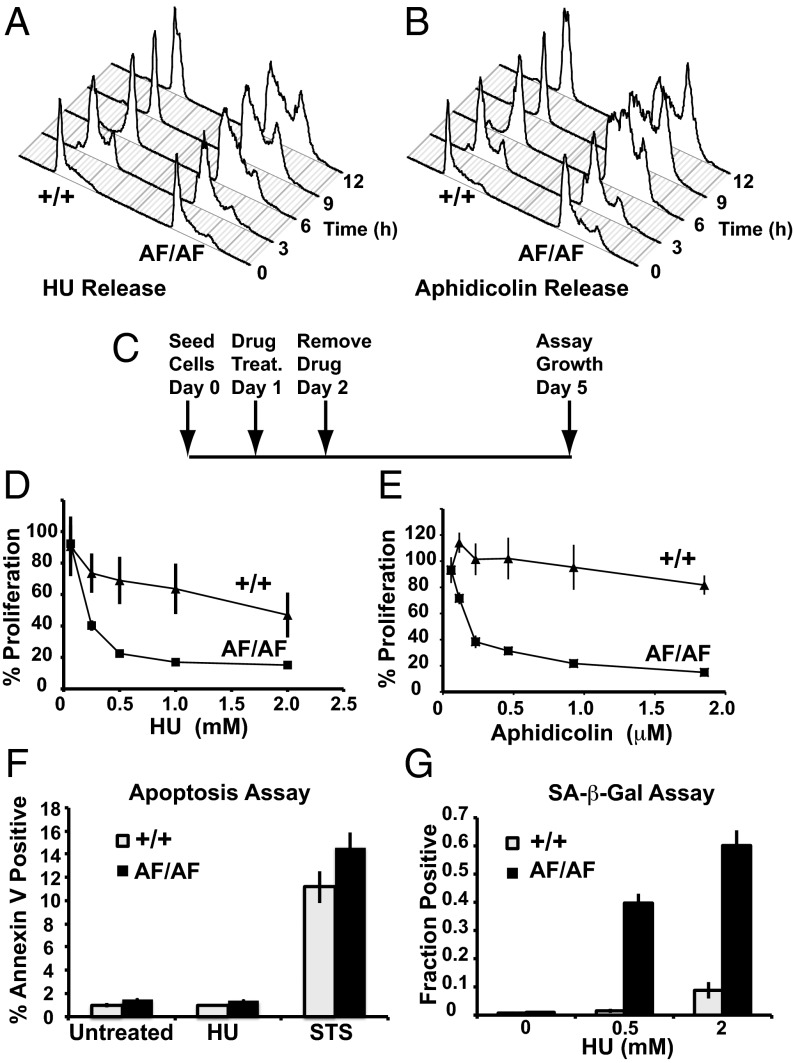
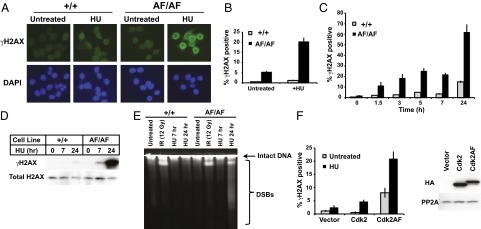
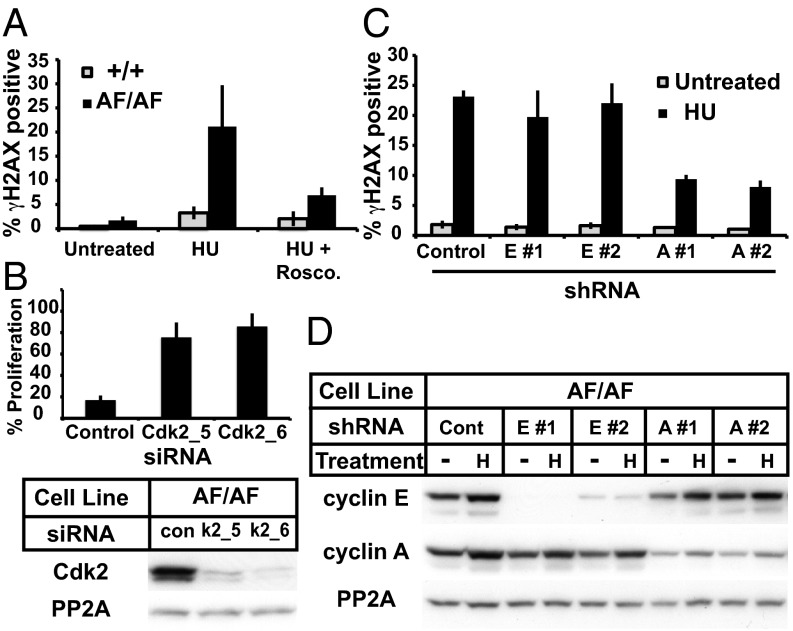
Cyclin-dependent kinases (Cdks) coordinate cell division, and their activities are tightly controlled. Phosphorylation of threonine 14 (T14) and tyrosine 15 (Y15) inhibits Cdks and regulates their activities in numerous physiologic contexts. Although the roles of Cdk1 inhibitory phosphorylation during mitosis are well described, studies of Cdk2 inhibitory phosphorylation during S phrase have largely been indirect. To specifically study the functions of Cdk2 inhibitory phosphorylation, we used gene targeting to make an endogenous Cdk2 knockin allele in human cells, termed Cdk2AF, which prevents Cdk2 T14 and Y15 phosphorylation. Cdk2AF caused premature S-phase entry, rapid cyclin E degradation, abnormal DNA replication, and genome instability. Cdk2AF cells also exhibited strikingly abnormal responses to replication stress, accumulated irreparable DNA damage, and permanently exited the cell cycle after transient exposure to S-phase inhibitors. Our results reveal the specific and essential roles of Cdk2 inhibitory phosphorylation in the successful execution of the replication stress checkpoint response and in maintaining genome integrity.
Keywords: Wee1; cyclin A.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Loss of cyclin-dependent kinase 2 (CDK2) inhibitory phosphorylation in a CDK2AF knock-in mouse causes misregulation of DNA replication and centrosome duplication.Mol Cell Biol. 2012 Apr;32(8):1421-32. doi: 10.1128/MCB.06721-11. Epub 2012 Feb 13. Mol Cell Biol. 2012. PMID: 22331465 Free PMC article.
-
Activation of Cdk2/Cyclin E complexes is dependent on the origin of replication licensing factor Cdc6 in mammalian cells.Cell Cycle. 2010 Nov 15;9(22):4533-41. doi: 10.4161/cc.9.22.13789. Epub 2010 Nov 15. Cell Cycle. 2010. PMID: 21088490
-
Identification and functional analysis of a novel cyclin e/cdk2 substrate ankrd17.J Biol Chem. 2009 Mar 20;284(12):7875-88. doi: 10.1074/jbc.M807827200. Epub 2009 Jan 16. J Biol Chem. 2009. PMID: 19150984 Free PMC article.
-
WEE1 kinase limits CDK activities to safeguard DNA replication and mitotic entry.Mutat Res. 2020 Jan-Apr;819-820:111694. doi: 10.1016/j.mrfmmm.2020.111694. Epub 2020 Feb 25. Mutat Res. 2020. PMID: 32120135 Review.
-
Cyclin-dependent kinases and S phase control in mammalian cells.Cell Cycle. 2003 Jul-Aug;2(4):316-24. Cell Cycle. 2003. PMID: 12851482 Review.
Cited by
-
Therapeutic targeting of replicative immortality.Semin Cancer Biol. 2015 Dec;35 Suppl(Suppl):S104-S128. doi: 10.1016/j.semcancer.2015.03.007. Epub 2015 Apr 11. Semin Cancer Biol. 2015. PMID: 25869441 Free PMC article. Review.
-
CDK4/6 inhibitor-induced bone marrow micronuclei might be caused by cell cycle arrest during erythropoiesis.Genes Environ. 2024 Feb 1;46(1):3. doi: 10.1186/s41021-024-00298-5. Genes Environ. 2024. PMID: 38303098 Free PMC article.
-
Emerging Roles for Ciz1 in Cell Cycle Regulation and as a Driver of Tumorigenesis.Biomolecules. 2016 Dec 27;7(1):1. doi: 10.3390/biom7010001. Biomolecules. 2016. PMID: 28036012 Free PMC article. Review.
-
Inhibition of the ATR-CHK1 Pathway in Ewing Sarcoma Cells Causes DNA Damage and Apoptosis via the CDK2-Mediated Degradation of RRM2.Mol Cancer Res. 2020 Jan;18(1):91-104. doi: 10.1158/1541-7786.MCR-19-0585. Epub 2019 Oct 24. Mol Cancer Res. 2020. PMID: 31649026 Free PMC article.
-
DNA replication stress: oncogenes in the spotlight.Genet Mol Biol. 2019 Dec 13;43(1 suppl 1):e20190138. doi: 10.1590/1678-4685GMB-2019-0138. eCollection 2019. Genet Mol Biol. 2019. PMID: 31930281 Free PMC article.
References
-
- Morgan DO. Principles of CDK regulation. Nature. 1995;374(6518):131–134. - PubMed
-
- Atherton-Fessler S, Hannig G, Piwnica-Worms H. Reversible tyrosine phosphorylation and cell cycle control. Semin Cell Biol. 1993;4(6):433–442. - PubMed
-
- Berry LD, Gould KL. Regulation of Cdc2 activity by phosphorylation at T14/Y15. Prog Cell Cycle Res. 1996;2:99–105. - PubMed
-
- Coleman TR, Dunphy WG. Cdc2 regulatory factors. Curr Opin Cell Biol. 1994;6(6):877–882. - PubMed
-
- Ferrell JE, Jr, et al. Simple, realistic models of complex biological processes: Positive feedback and bistability in a cell fate switch and a cell cycle oscillator. FEBS Lett. 2009;583(24):3999–4005. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
