

The EGF repeat-specific O-GlcNAc-transferase Eogt interacts with notch signaling and pyrimidine metabolism pathways in Drosophila
- PMID: 23671640
- PMCID: PMC3650022
- DOI: 10.1371/journal.pone.0062835
The EGF repeat-specific O-GlcNAc-transferase Eogt interacts with notch signaling and pyrimidine metabolism pathways in Drosophila
Abstract
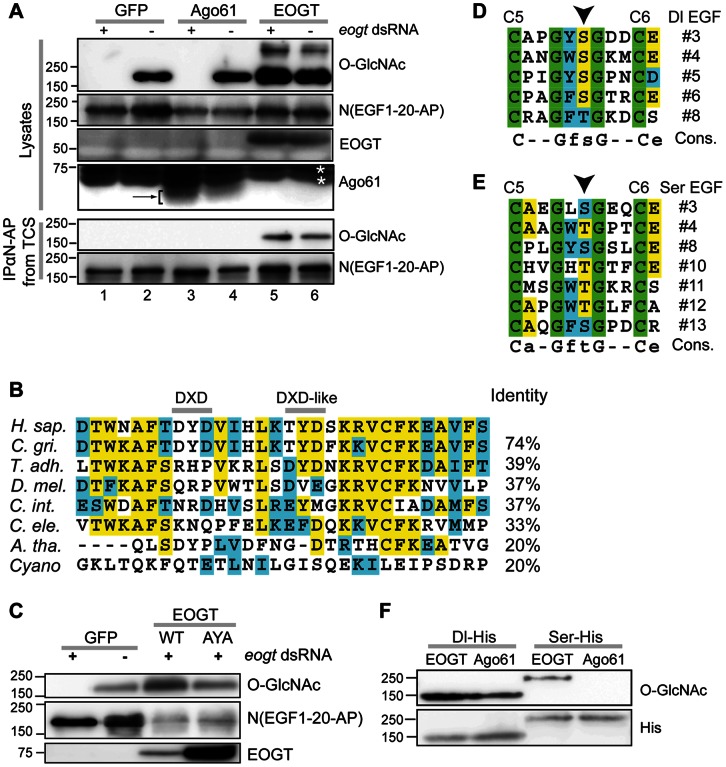
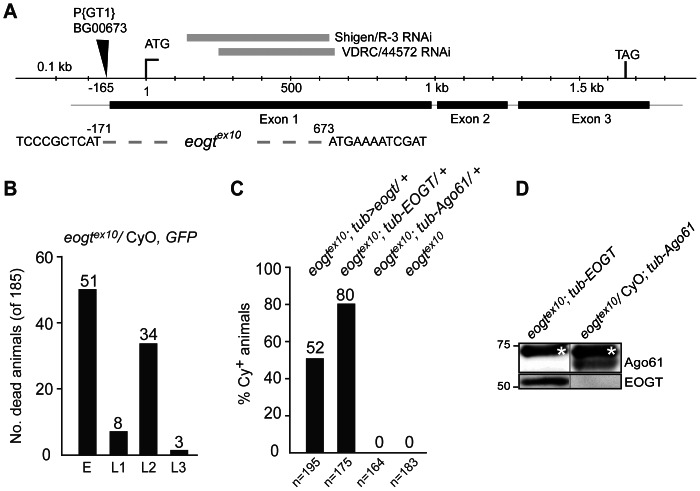
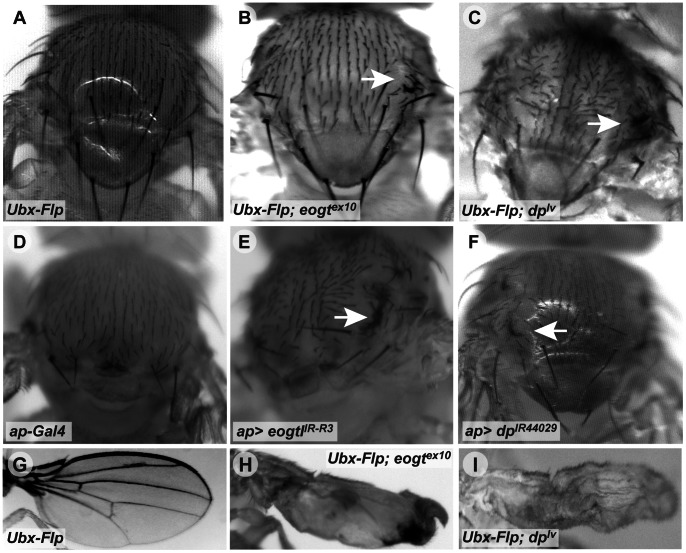
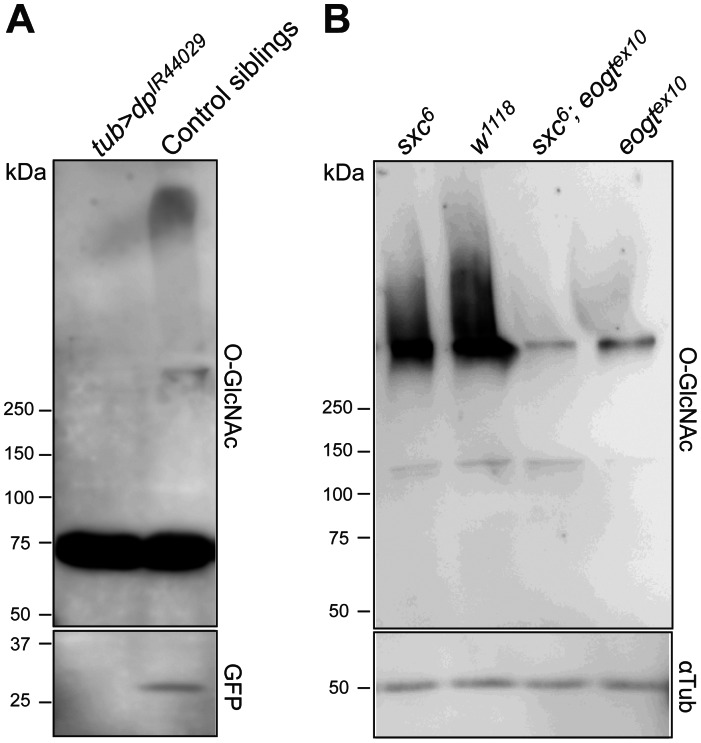
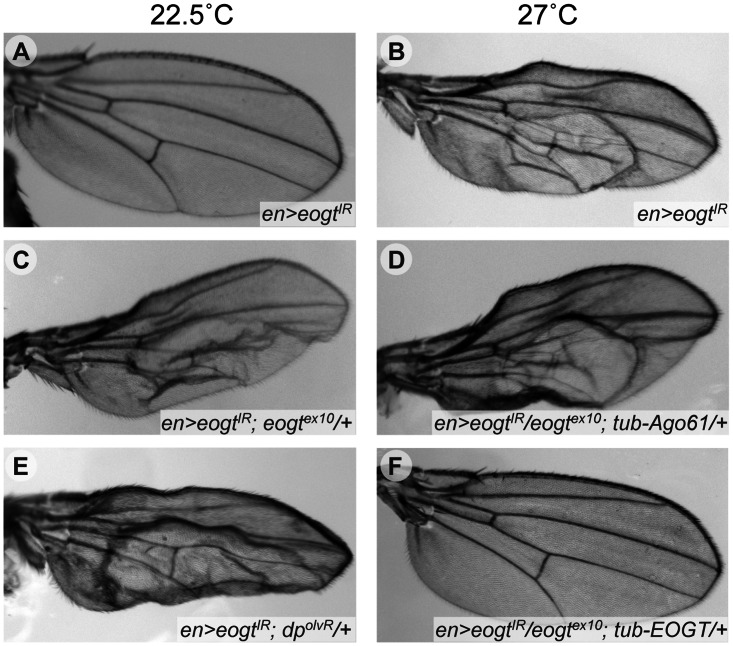
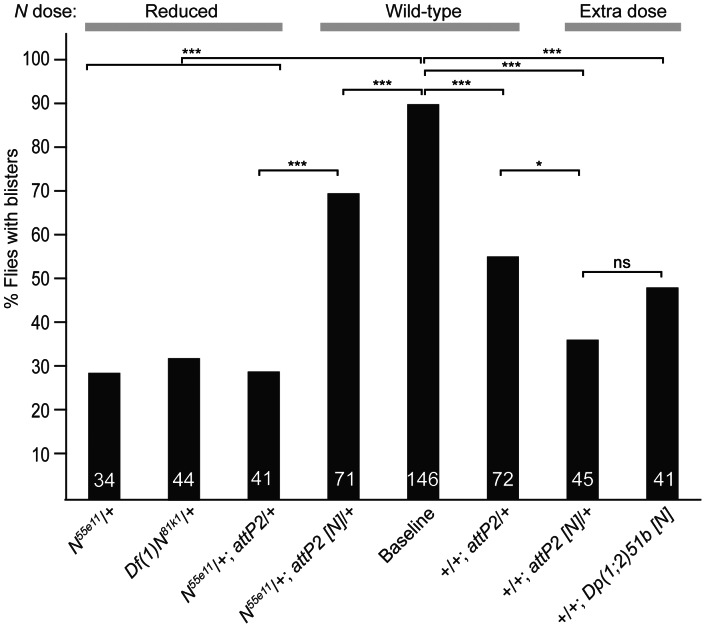
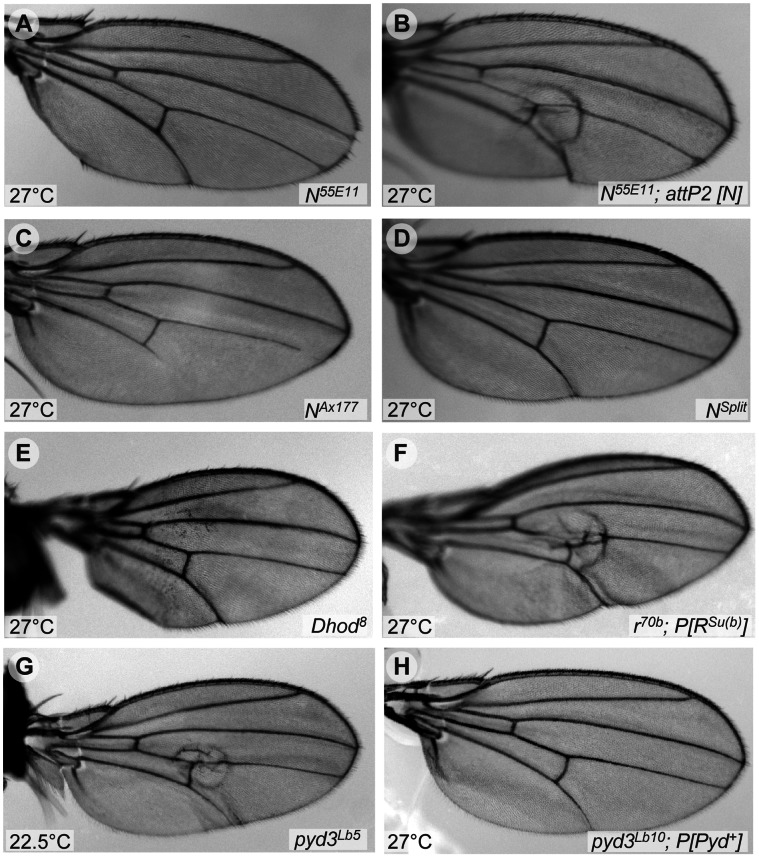
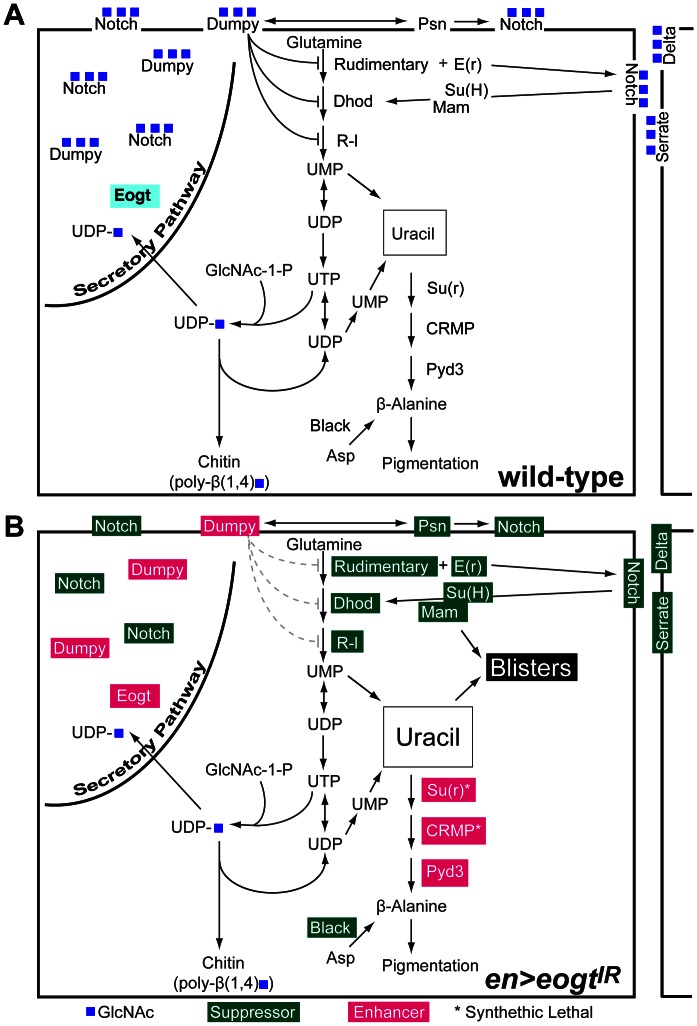
The O-GlcNAc transferase Eogt modifies EGF repeats in proteins that transit the secretory pathway, including Dumpy and Notch. In this paper, we show that the Notch ligands Delta and Serrate are also substrates of Eogt, that mutation of a putative UDP-GlcNAc binding DXD motif greatly reduces enzyme activity, and that Eogt and the cytoplasmic O-GlcNAc transferase Ogt have distinct substrates in Drosophila larvae. Loss of Eogt is larval lethal and disrupts Dumpy functions, but does not obviously perturb Notch signaling. To identify novel genetic interactions with eogt, we investigated dominant modification of wing blister formation caused by knock-down of eogt. Unexpectedly, heterozygosity for several members of the canonical Notch signaling pathway suppressed wing blister formation. And importantly, extensive genetic interactions with mutants in pyrimidine metabolism were identified. Removal of pyrimidine synthesis alleles suppressed wing blister formation, while removal of uracil catabolism alleles was synthetic lethal with eogt knock-down. Therefore, Eogt may regulate protein functions by O-GlcNAc modification of their EGF repeats, and cellular metabolism by affecting pyrimidine synthesis and catabolism. We propose that eogt knock-down in the wing leads to metabolic and signaling perturbations that increase cytosolic uracil levels, thereby causing wing blister formation.
Conflict of interest statement
Figures
Similar articles
-
Impaired O-linked N-acetylglucosaminylation in the endoplasmic reticulum by mutated epidermal growth factor (EGF) domain-specific O-linked N-acetylglucosamine transferase found in Adams-Oliver syndrome.J Biol Chem. 2015 Jan 23;290(4):2137-49. doi: 10.1074/jbc.M114.598821. Epub 2014 Dec 8. J Biol Chem. 2015. PMID: 25488668 Free PMC article.
-
O-linked-N-acetylglucosamine modification of mammalian Notch receptors by an atypical O-GlcNAc transferase Eogt1.Biochem Biophys Res Commun. 2012 Mar 2;419(1):14-9. doi: 10.1016/j.bbrc.2012.01.098. Epub 2012 Jan 28. Biochem Biophys Res Commun. 2012. PMID: 22310717
-
EOGT and O-GlcNAc on secreted and membrane proteins.Biochem Soc Trans. 2017 Apr 15;45(2):401-408. doi: 10.1042/BST20160165. Biochem Soc Trans. 2017. PMID: 28408480 Free PMC article. Review.
-
N-Glycans on EGF domain-specific O-GlcNAc transferase (EOGT) facilitate EOGT maturation and peripheral endoplasmic reticulum localization.J Biol Chem. 2020 Jun 19;295(25):8560-8574. doi: 10.1074/jbc.RA119.012280. Epub 2020 May 6. J Biol Chem. 2020. PMID: 32376684 Free PMC article.
-
N-acetylglucosamine modification in the lumen of the endoplasmic reticulum.Biochim Biophys Acta. 2015 Jun;1850(6):1319-24. doi: 10.1016/j.bbagen.2015.03.003. Epub 2015 Mar 17. Biochim Biophys Acta. 2015. PMID: 25791024 Review.
Cited by
-
Proteomic Analysis of Pig (Sus scrofa) Olfactory Soluble Proteome Reveals O-Linked-N-Acetylglucosaminylation of Secreted Odorant-Binding Proteins.Front Endocrinol (Lausanne). 2014 Dec 5;5:202. doi: 10.3389/fendo.2014.00202. eCollection 2014. Front Endocrinol (Lausanne). 2014. PMID: 25538681 Free PMC article.
-
Cell Communications among Microorganisms, Plants, and Animals: Origin, Evolution, and Interplays.Int J Mol Sci. 2020 Oct 28;21(21):8052. doi: 10.3390/ijms21218052. Int J Mol Sci. 2020. PMID: 33126770 Free PMC article. Review.
-
Structural Divergence in O-GlcNAc Glycans Displayed on Epidermal Growth Factor-like Repeats of Mammalian Notch1.Molecules. 2018 Jul 17;23(7):1745. doi: 10.3390/molecules23071745. Molecules. 2018. PMID: 30018219 Free PMC article.
-
Impaired O-linked N-acetylglucosaminylation in the endoplasmic reticulum by mutated epidermal growth factor (EGF) domain-specific O-linked N-acetylglucosamine transferase found in Adams-Oliver syndrome.J Biol Chem. 2015 Jan 23;290(4):2137-49. doi: 10.1074/jbc.M114.598821. Epub 2014 Dec 8. J Biol Chem. 2015. PMID: 25488668 Free PMC article.
-
Structure, function, and pathology of protein O-glucosyltransferases.Cell Death Dis. 2021 Jan 12;12(1):71. doi: 10.1038/s41419-020-03314-y. Cell Death Dis. 2021. PMID: 33436558 Free PMC article. Review.
References
-
- Varki A, Cummings RD, Esko JD, Freeze HH, Stanley P, et al... (2009) Essentials of Glycobiology; Varki A, Cummings RD, Esko JD, Freeze HH, Stanley P et al., editors. New York: Cold Spring Harbor laboratory Press. - PubMed
-
- Moloney DJ, Panin VM, Johnston SH, Chen J, Shao L, et al. (2000) Fringe is a glycosyltransferase that modifies Notch. Nature 406: 369–375. - PubMed
-
- Bruckner K, Perez L, Clausen H, Cohen S (2000) Glycosyltransferase activity of Fringe modulates Notch-Delta interactions. Nature 406: 411–415. - PubMed
-
- Irvine KD (1999) Fringe, Notch, and making developmental boundaries. Curr Opin Genet Dev 9: 434–441. - PubMed
-
- Stanley P, Okajima T (2010) Roles of glycosylation in Notch signaling. Curr Top Dev Biol 92: 131–164. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
