

Regulation of de novo ceramide synthesis: the role of dihydroceramide desaturase and transcriptional factors NFATC and Hand2 in the hypoxic mouse heart
- PMID: 23672204
- PMCID: PMC3665309
- DOI: 10.1089/dna.2013.1993
Regulation of de novo ceramide synthesis: the role of dihydroceramide desaturase and transcriptional factors NFATC and Hand2 in the hypoxic mouse heart
Abstract
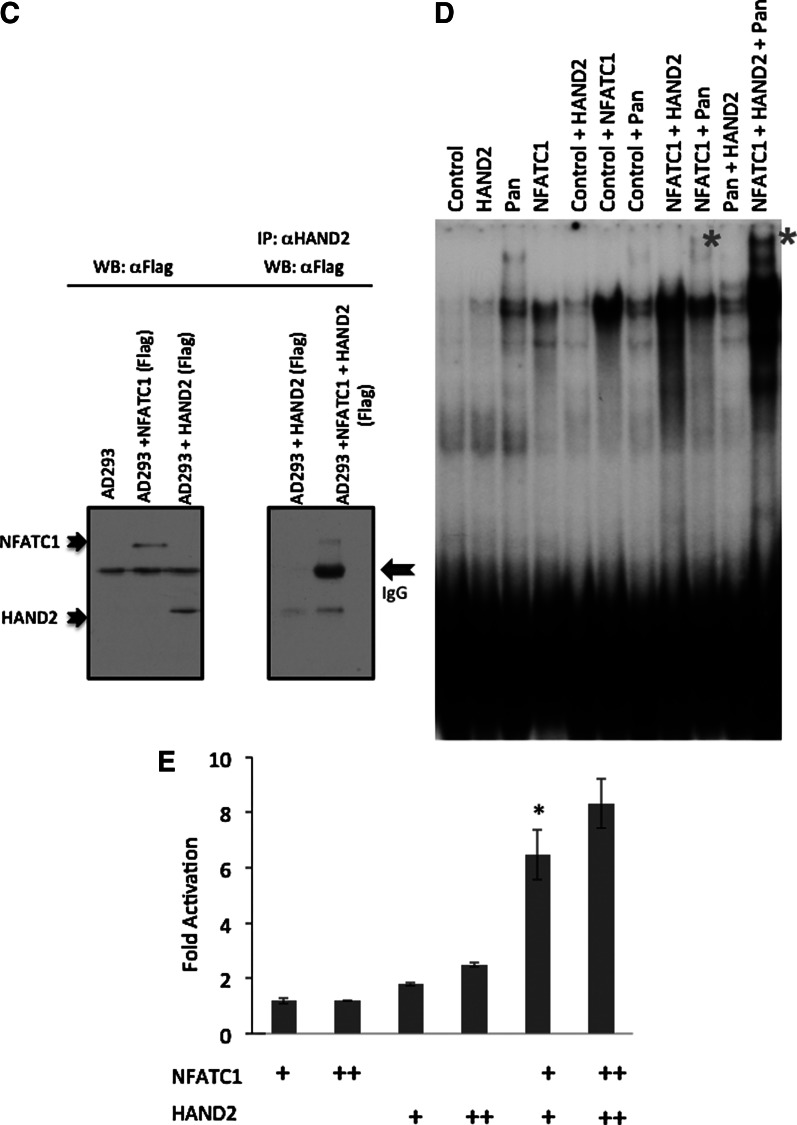
We have previously shown that ceramide, a proapoptotic molecule decreases in the mouse heart as it adapts to hypoxia. We have also shown that its precursor, dihydroceramide, accumulates with hypoxia. This implicates the enzyme dihydroceramide desaturase (DHC-DS), which converts dihydroceramide to ceramide, in a potential regulatory checkpoint in cardiomyocytes. We hypothesised that the regulation of de novo ceramide synthesis plays an important role in the cardiomyocyte adaptation to hypoxia. We used an established mouse model to induce acute and chronic hypoxia. Cardiac tissues were extracted and quantitative real-time polymerase chain reaction (qRT-PCR) was used to evaluate the expression levels of DHC-DS. Electrophoretic Mobility Shift Assays (EMSAs) and qRT-PCR were used to evaluate the activity and expression levels of an array of transcription factors that might regulate DEGS1 gene expression. We demonstrated that DEGS1 mRNA levels decrease with time in hypoxic mice concurrent with the decrease in HAND2 transcripts. Interestingly, the DEGS1 promoter harbors overlapping sites for Hand2 and Nuclear Factor of Activated T-cells (NFATC) transcription factors. We have demonstrated a physical interaction between NFATC1 and the E-Box proteins with EMSA and coimmunoprecipitation assays. The regulation of de novo ceramide synthesis in response to hypoxia and this newly described interaction between E-box and NFATC transcription factors will pave the way to identify new pathways in the adaptation of the cardiomyocyte to stress. The elucidation of these pathways will in the long-term provide insights into potential targets for novel therapeutic regimens.
Figures





Similar articles
-
Dihydroceramide desaturase 1, the gatekeeper of ceramide induced lipotoxicity.Biochim Biophys Acta. 2015 Jan;1851(1):40-50. doi: 10.1016/j.bbalip.2014.09.021. Epub 2014 Oct 2. Biochim Biophys Acta. 2015. PMID: 25283058 Review.
-
Dihydroceramide Desaturase Functions as an Inducer and Rectifier of Apoptosis: Effect of Retinol Derivatives, Antioxidants and Phenolic Compounds.Cell Biochem Biophys. 2021 Sep;79(3):461-475. doi: 10.1007/s12013-021-00990-1. Epub 2021 May 15. Cell Biochem Biophys. 2021. PMID: 33991313 Free PMC article.
-
MicroRNA-92b-3p suppresses angiotensin II-induced cardiomyocyte hypertrophy via targeting HAND2.Life Sci. 2019 Sep 1;232:116635. doi: 10.1016/j.lfs.2019.116635. Epub 2019 Jul 5. Life Sci. 2019. PMID: 31283925
-
Modulation of total ceramide and constituent ceramide species in the acutely and chronically hypoxic mouse heart at different ages.Prostaglandins Other Lipid Mediat. 2008 Jun;86(1-4):49-55. doi: 10.1016/j.prostaglandins.2008.02.003. Epub 2008 Mar 4. Prostaglandins Other Lipid Mediat. 2008. PMID: 18400538
-
New role for ceramide in hypoxia and insulin resistance.World J Gastroenterol. 2020 May 14;26(18):2177-2186. doi: 10.3748/wjg.v26.i18.2177. World J Gastroenterol. 2020. PMID: 32476784 Free PMC article. Review.
Cited by
-
Potential Drug Targets for Ceramide Metabolism in Cardiovascular Disease.J Cardiovasc Dev Dis. 2022 Dec 2;9(12):434. doi: 10.3390/jcdd9120434. J Cardiovasc Dev Dis. 2022. PMID: 36547431 Free PMC article. Review.
-
Dihydroceramides: From Bit Players to Lead Actors.J Biol Chem. 2015 Jun 19;290(25):15371-15379. doi: 10.1074/jbc.R115.653204. Epub 2015 May 6. J Biol Chem. 2015. PMID: 25947377 Free PMC article. Review.
-
Regulation of ceramide channel formation and disassembly: Insights on the initiation of apoptosis.Saudi J Biol Sci. 2015 Nov;22(6):760-72. doi: 10.1016/j.sjbs.2015.03.005. Epub 2015 Mar 22. Saudi J Biol Sci. 2015. PMID: 26587005 Free PMC article.
-
Sphingolipid abnormalities in cancer multidrug resistance: Chicken or egg?Cell Signal. 2017 Oct;38:134-145. doi: 10.1016/j.cellsig.2017.06.017. Epub 2017 Jul 4. Cell Signal. 2017. PMID: 28687494 Free PMC article. Review.
-
Mysterious sphingolipids: metabolic interrelationships at the center of pathophysiology.Front Physiol. 2024 Jan 3;14:1229108. doi: 10.3389/fphys.2023.1229108. eCollection 2023. Front Physiol. 2024. PMID: 38235387 Free PMC article. Review.
References
-
- Adrogue J.V. Sharma S. Ngumbela K. Essop M.F. Taegtmeyer H. Acclimatization to chronic hypobaric hypoxia is associated with a differential transcriptional profile between the right and left ventricle. Mol Cell Biochem. 2005;278:71–78. - PubMed
-
- Bruneau B.G. Nemer G. Schmitt J.P. Charron F. Robitaille L. Caron S. Conner D.A. Gessler M. Nemer M. Seidman C.E., et al. A murine model of Holt-Oram syndrome defines roles of the T-box transcription factor Tbx5 in cardiogenesis and disease. Cell. 2001;106:709–721. - PubMed
-
- Geley S. Hartmann B.L. Kofler R. Ceramides induce a form of apoptosis in human acute lymphoblastic leukemia cells that is inhibited by Bcl-2, but not by CrmA. FEBS Lett. 1997;400:15–18. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
