

Muscle-specific Pikfyve gene disruption causes glucose intolerance, insulin resistance, adiposity, and hyperinsulinemia but not muscle fiber-type switching
- PMID: 23673157
- PMCID: PMC3725567
- DOI: 10.1152/ajpendo.00030.2013
Muscle-specific Pikfyve gene disruption causes glucose intolerance, insulin resistance, adiposity, and hyperinsulinemia but not muscle fiber-type switching
Abstract
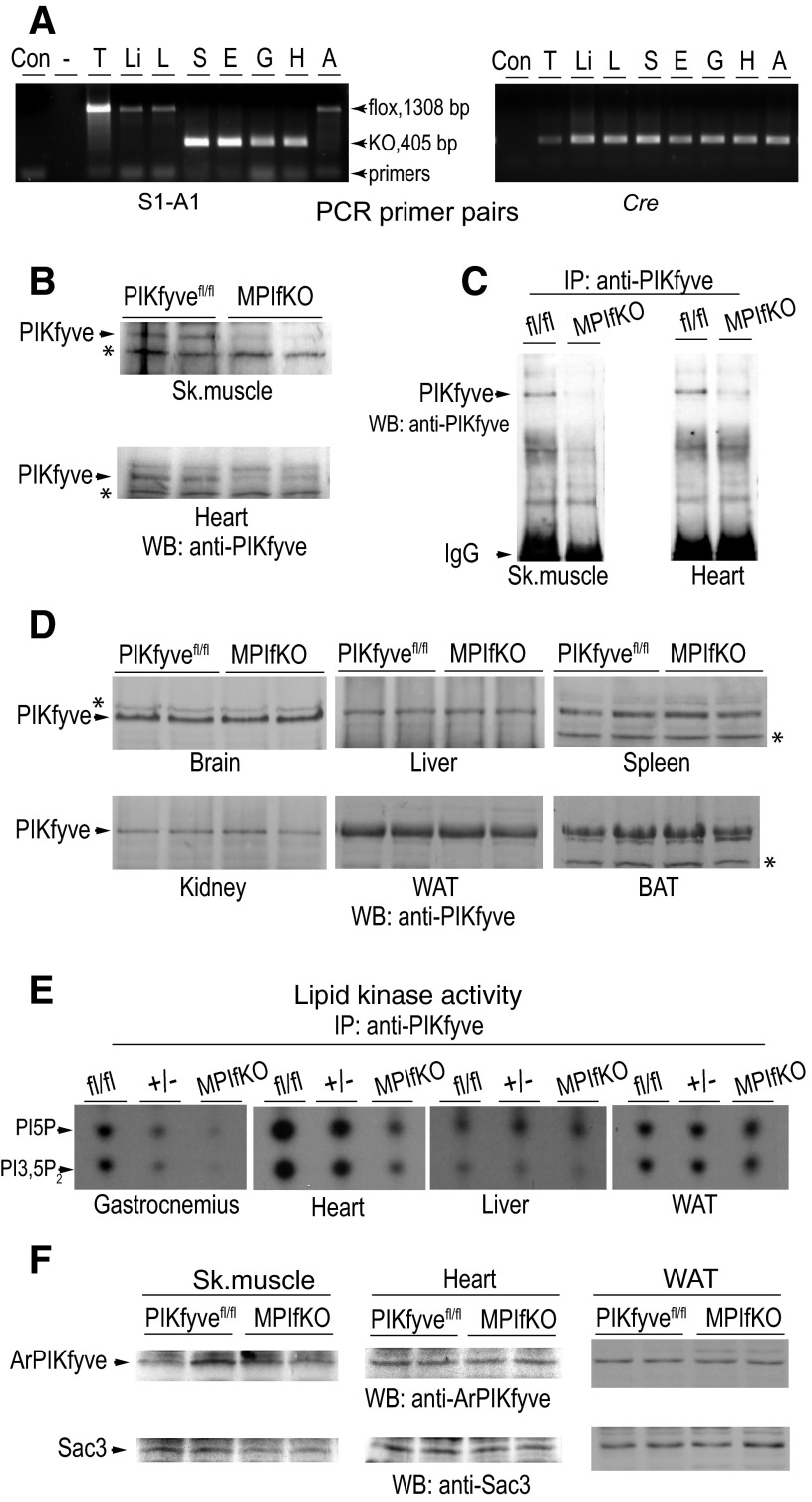
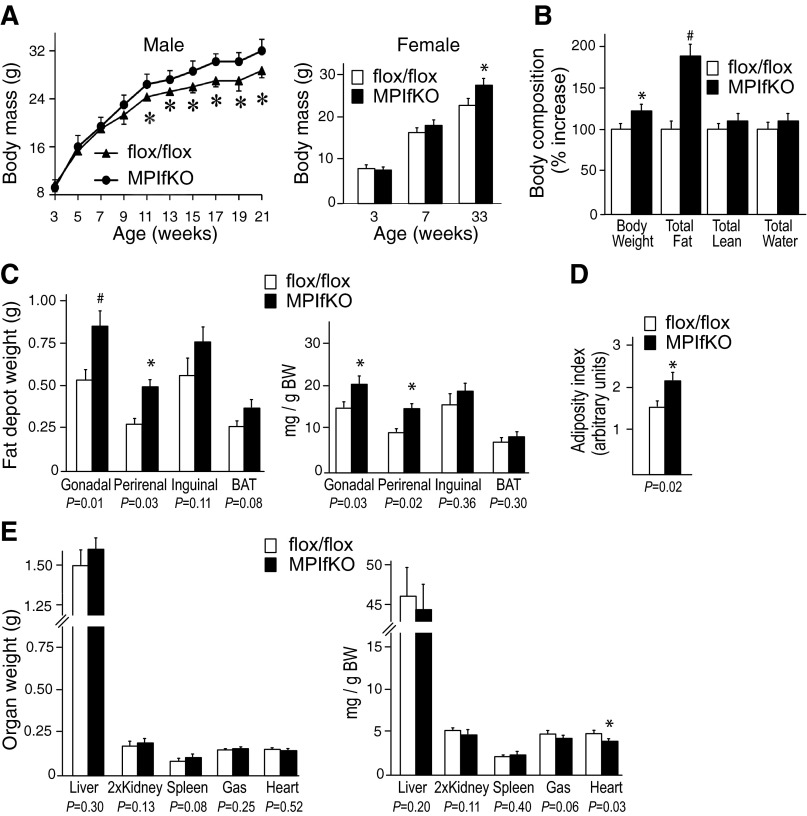
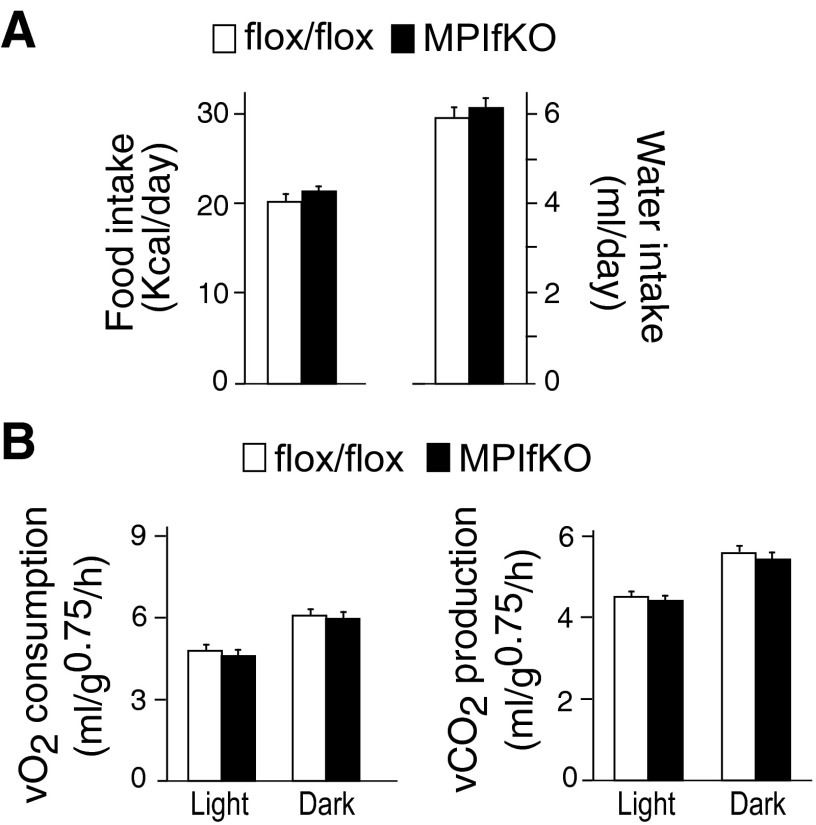
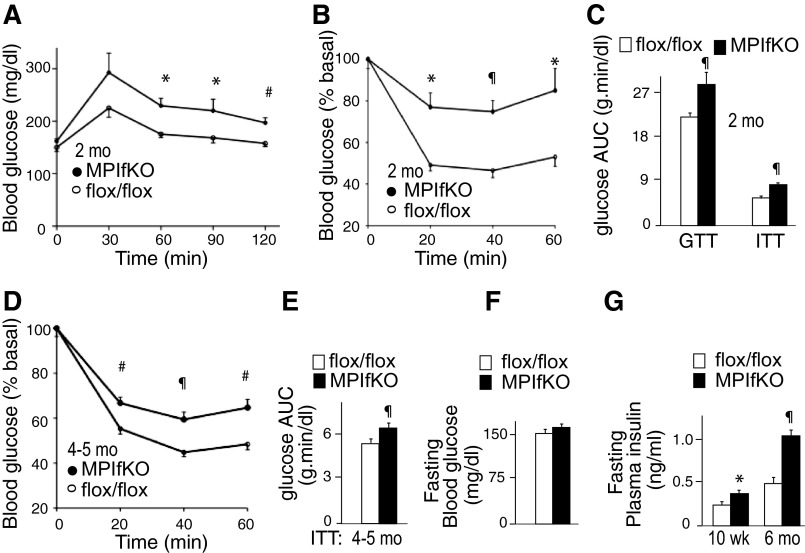
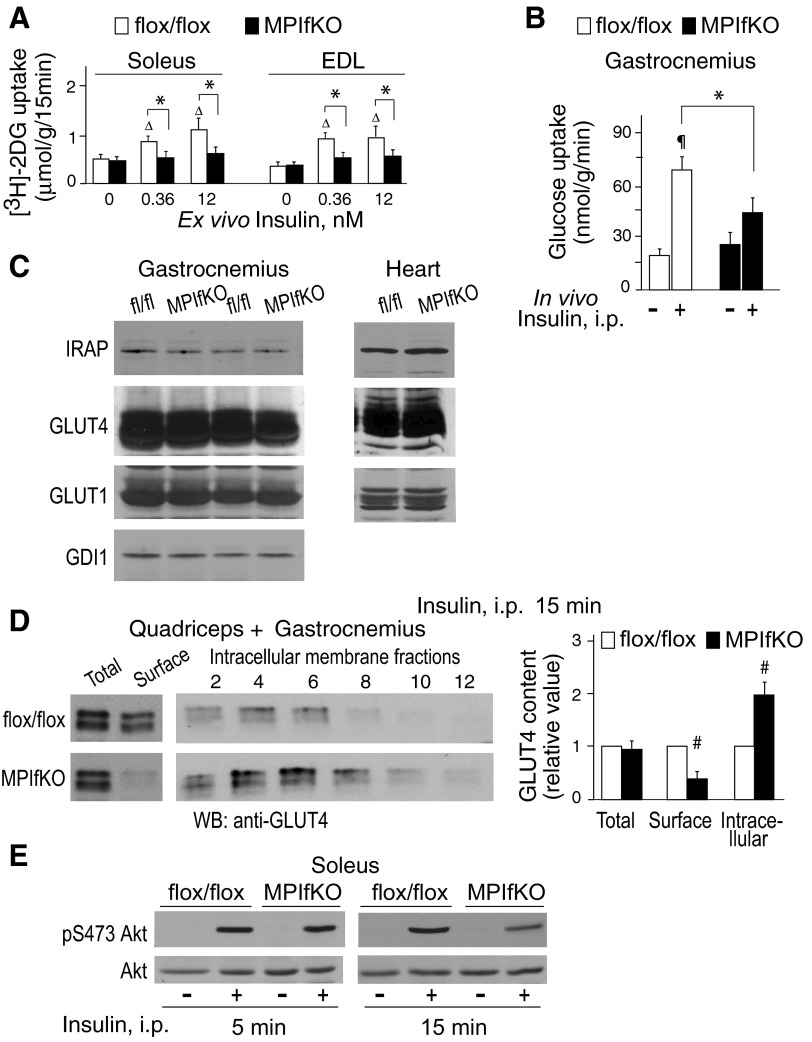
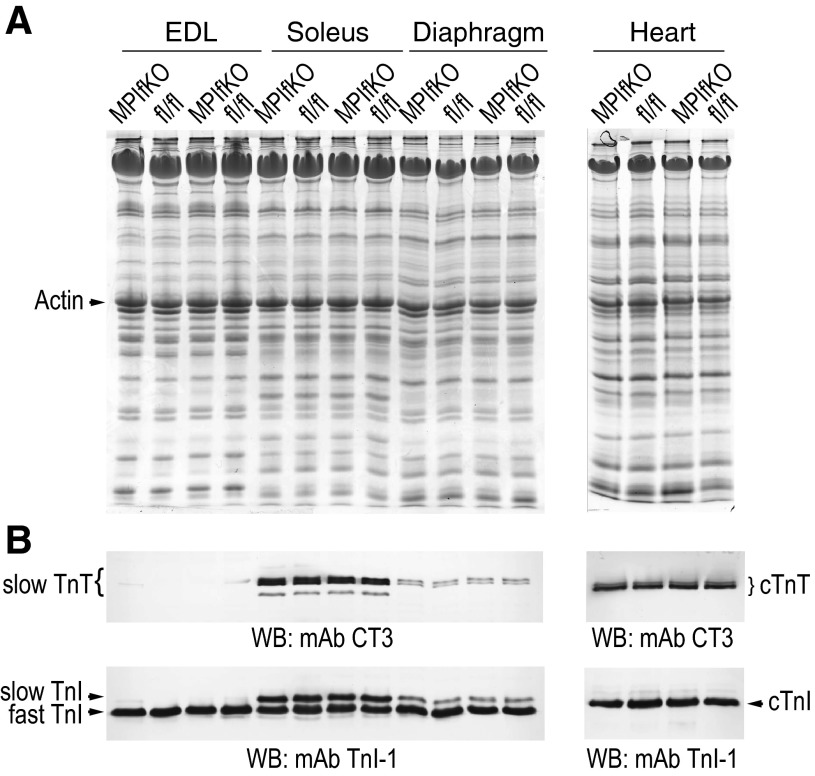
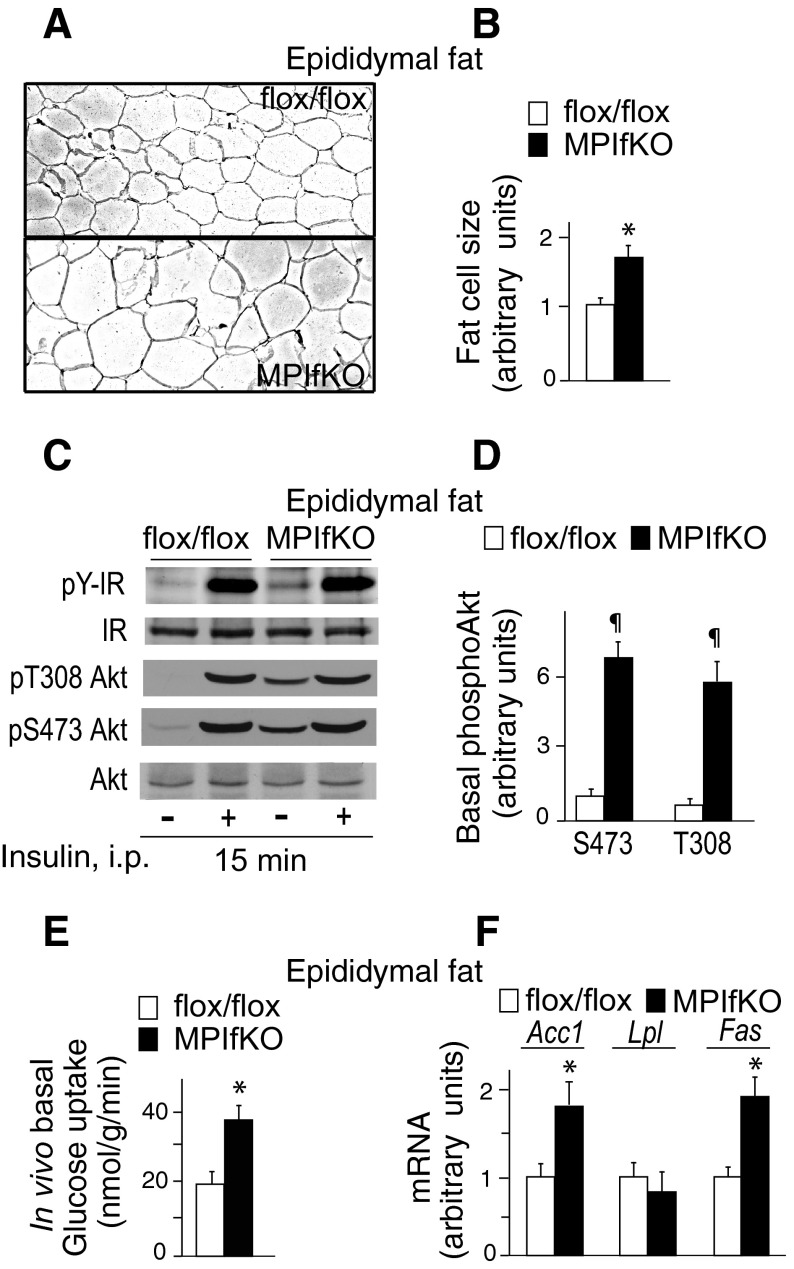
The evolutionarily conserved kinase PIKfyve that synthesizes PtdIns5P and PtdIns(3,5)P₂ has been implicated in insulin-regulated GLUT4 translocation/glucose entry in 3T3-L1 adipocytes. To decipher PIKfyve's role in muscle and systemic glucose metabolism, here we have developed a novel mouse model with Pikfyve gene disruption in striated muscle (MPIfKO). These mice exhibited systemic glucose intolerance and insulin resistance at an early age but had unaltered muscle mass or proportion of slow/fast-twitch muscle fibers. Insulin stimulation of in vivo or ex vivo glucose uptake and GLUT4 surface translocation was severely blunted in skeletal muscle. These changes were associated with premature attenuation of Akt phosphorylation in response to in vivo insulin, as tested in young mice. Starting at 10-11 wk of age, MPIfKO mice progressively accumulated greater body weight and fat mass. Despite increased adiposity, serum free fatty acid and triglyceride levels were normal until adulthood. Together with the undetectable lipid accumulation in liver, these data suggest that lipotoxicity and muscle fiber switching do not contribute to muscle insulin resistance in MPIfKO mice. Furthermore, the 80% increase in total fat mass resulted from increased fat cell size rather than altered fat cell number. The observed profound hyperinsulinemia combined with the documented increases in constitutive Akt activation, in vivo glucose uptake, and gene expression of key enzymes for fatty acid biosynthesis in MPIfKO fat tissue suggest that the latter is being sensitized for de novo lipid anabolism. Our data provide the first in vivo evidence that PIKfyve is essential for systemic glucose homeostasis and insulin-regulated glucose uptake/GLUT4 translocation in skeletal muscle.
Keywords: PIKfyve metabolic functions; PIKfyve muscle knockout; insulin resistance; insulin-regulated glucose transporter 4 translocation; muscle glucose uptake; prediabetes; systemic glucose homeostasis.
Figures
References
-
- Abdul-Ghani MA, DeFronzo RA. Pathophysiology of prediabetes. Curr Diab Rep 9: 193–199, 2009 - PubMed
-
- Biddinger SB, Kahn CR. From mice to men: insights into the insulin resistance syndromes. Annu Rev Physiol 68: 123–158, 2006 - PubMed
-
- Björnholm M, Zierath JR. Insulin signal transduction in human skeletal muscle: identifying the defects in Type II diabetes. Biochem Soc Trans 33: 354–357, 2005 - PubMed
-
- Bruning JC, Michael MD, Winnay JN, Hayashi T, Horsch D, Accili D, Goodyear LJ, Kahn CR. A muscle-specific insulin receptor knockout exhibits features of the metabolic syndrome of NIDDM without altering glucose tolerance. Mol Cell 2: 559–569, 1998 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases