

Long-term fluoxetine treatment induces input-specific LTP and LTD impairment and structural plasticity in the CA1 hippocampal subfield
- PMID: 23675317
- PMCID: PMC3648695
- DOI: 10.3389/fncel.2013.00066
Long-term fluoxetine treatment induces input-specific LTP and LTD impairment and structural plasticity in the CA1 hippocampal subfield
Abstract
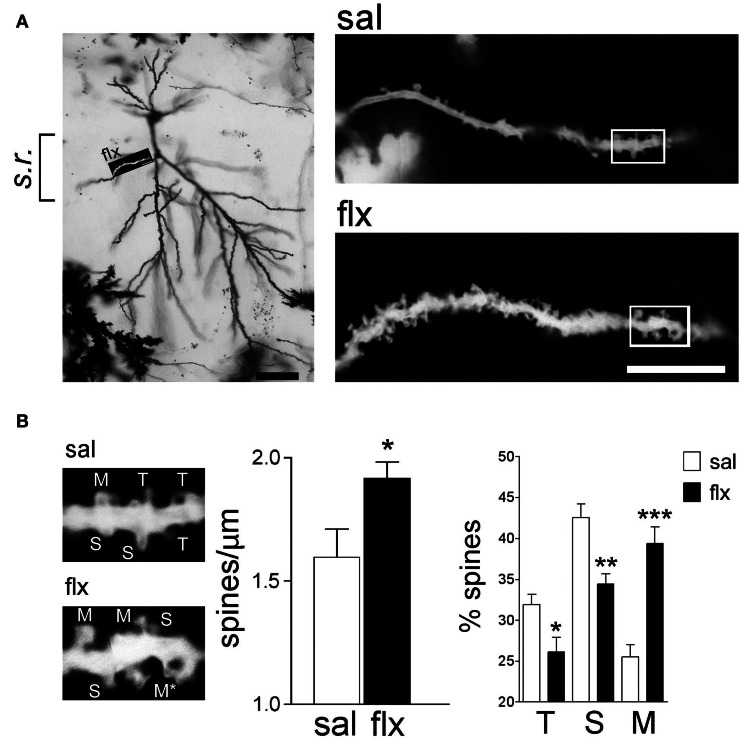
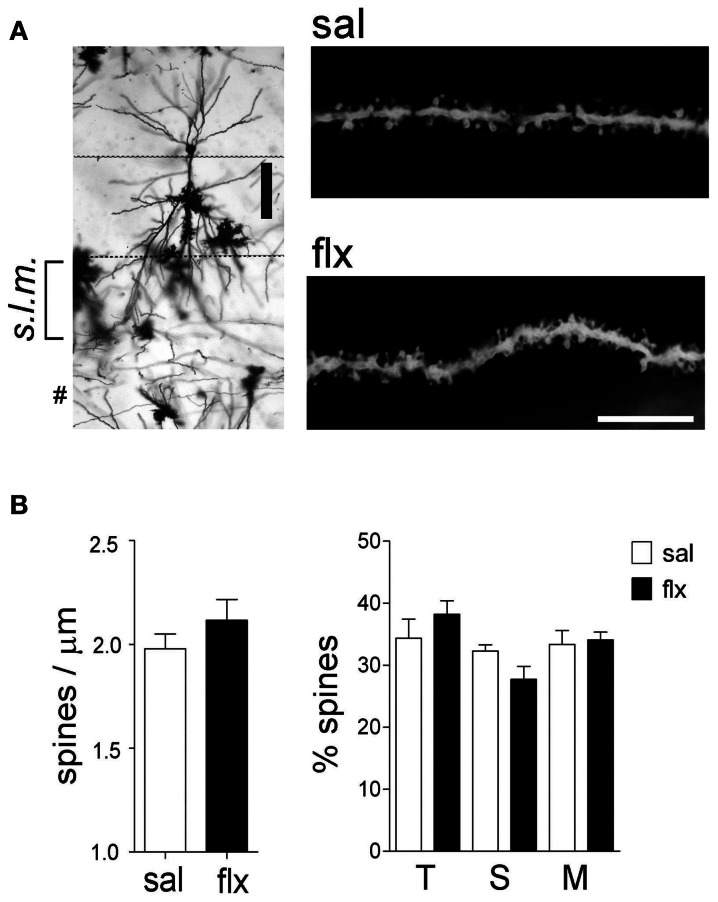
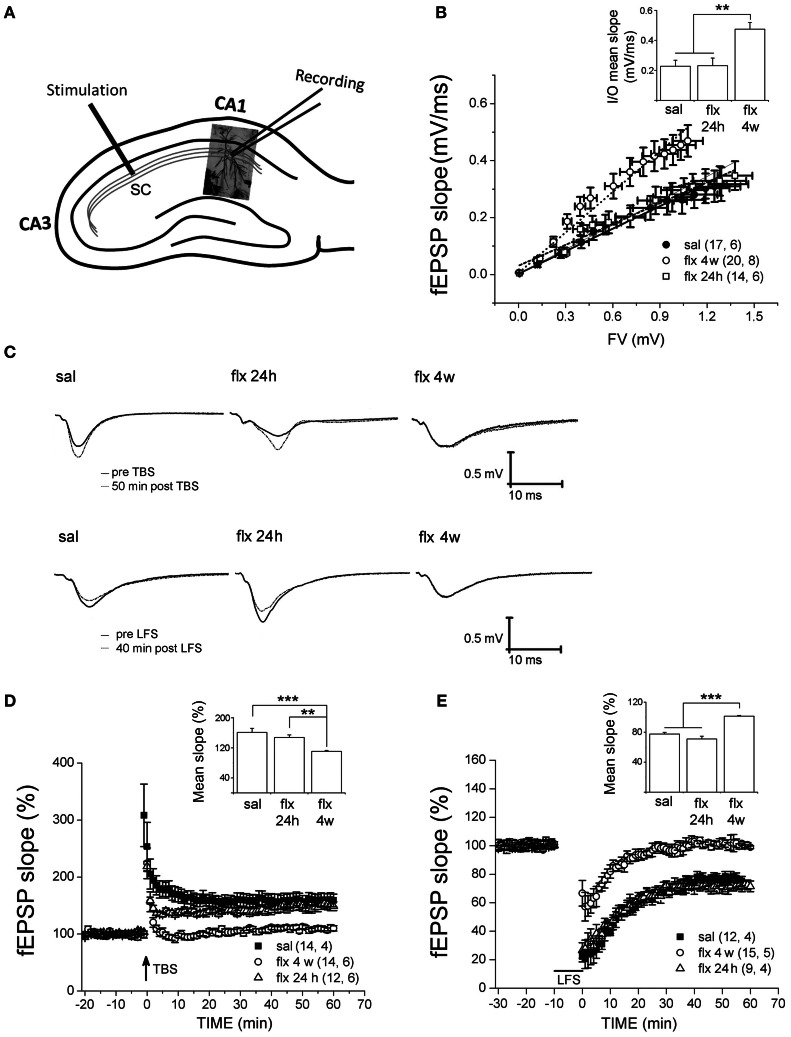
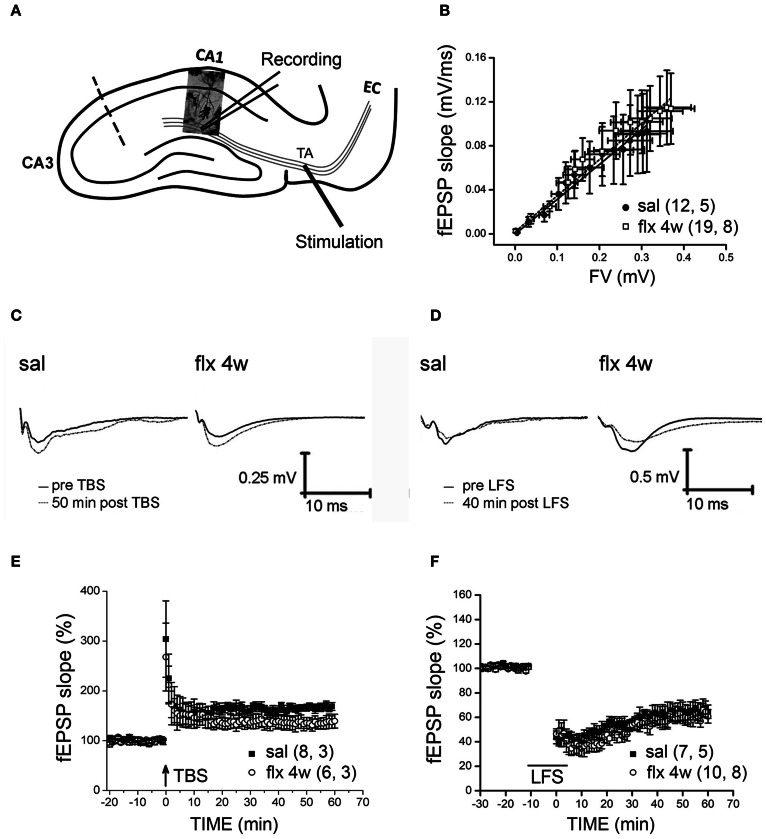
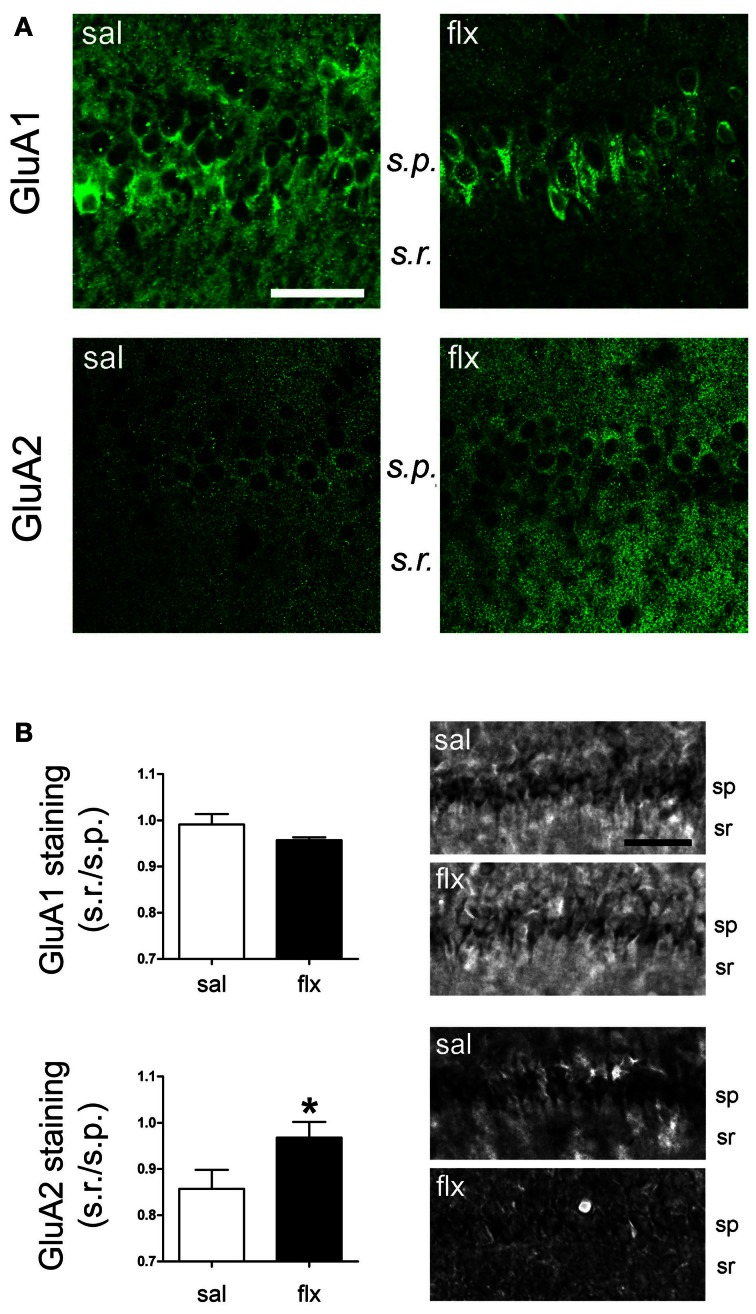
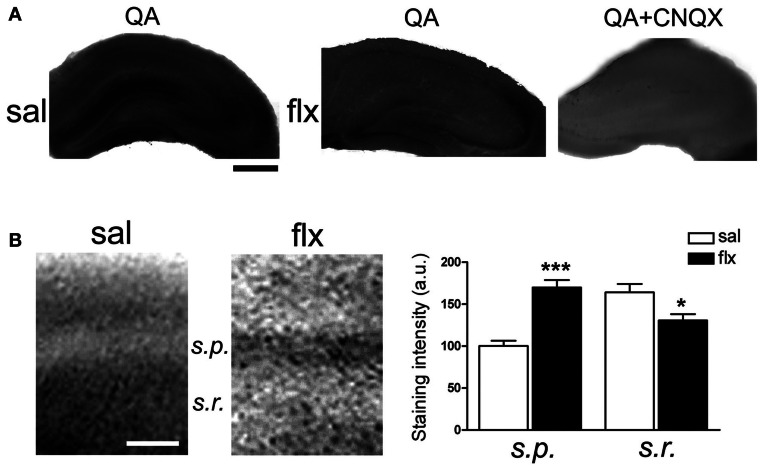
Antidepressant drugs are usually administered for several weeks for the treatment of major depressive disorder. However, they are also prescribed in several additional psychiatric conditions as well as during long-term maintenance treatments. Antidepressants induce adaptive changes in several forebrain structures which include modifications at glutamatergic synapses. We recently found that repetitive administration of the selective serotonin reuptake inhibitor (SSRI) fluoxetine to naïve adult male rats induced an increase of mature, mushroom-type dendritic spines in several forebrain regions. This was associated with an increase of GluA2-containing α-amino-3-hydroxy-5-methylisoxazole-4-propionate receptors (AMPA-Rs) in telencephalic postsynaptic densities. To unravel the functional significance of such a synaptic re-arrangement, we focused on glutamate neurotransmission in the hippocampus. We evaluated the effect of four weeks of 0.7 mg/kg fluoxetine on long-term potentiation (LTP) and long-term depression (LTD) in the CA1 hippocampal subfield. Recordings in hippocampal slices revealed profound deficits in LTP and LTD at Schaffer collateral-CA1 synapses associated to increased spine density and enhanced presence of mushroom-type spines, as revealed by Golgi staining. However, the same treatment had neither an effect on spine morphology, nor on LTP and LTD at perforant path-CA1 synapses. Cobalt staining and immunohistochemical experiments revealed decreased AMPA-R Ca(2+) permeability in the stratum radiatum (s.r.) together with increased GluA2-containing Ca(2+) impermeable AMPA-Rs. Therefore, 4 weeks of fluoxetine treatment promoted structural and functional adaptations in CA1 neurons in a pathway-specific manner that were selectively associated with impairment of activity-dependent plasticity at Schaffer collateral-CA1 synapses.
Keywords: LTD; LTP; antidepressants; dendritic spines; glutamate receptors.
Figures
References
-
- Ampuero E., Dagnino-Subiabre A., Sandoval R., Zepeda-Carreno R., Sandoval S., Viedma A., et al. (2007). Status epilepticus induces region-specific changes in dendritic spines, dendritic length and TrkB protein content of rat brain cortex. Brain Res. 1150, 225–238 10.1016/j.brainres.2007.02.089 - DOI - PubMed
-
- Ampuero E., Rubio F. J., Falcon R., Sandoval M., Diaz-Veliz G., Gonzalez R. E., et al. (2010). Chronic fluoxetine treatment induces structural plasticity and selective changes in glutamate receptor subunits in the rat cerebral cortex. Neuroscience 169, 98–108 10.1016/j.neuroscience.2010.04.035 - DOI - PubMed
-
- Andrade R., Chaput Y. (1991). 5-Hydroxytryptamine4-like receptors mediate the slow excitatory response to serotonin in the rat hippocampus. J. Pharmacol. Exp. Ther. 257, 930–937 - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous