

Visually driven chaining of elementary swim patterns into a goal-directed motor sequence: a virtual reality study of zebrafish prey capture
- PMID: 23675322
- PMCID: PMC3650304
- DOI: 10.3389/fncir.2013.00086
Visually driven chaining of elementary swim patterns into a goal-directed motor sequence: a virtual reality study of zebrafish prey capture
Abstract
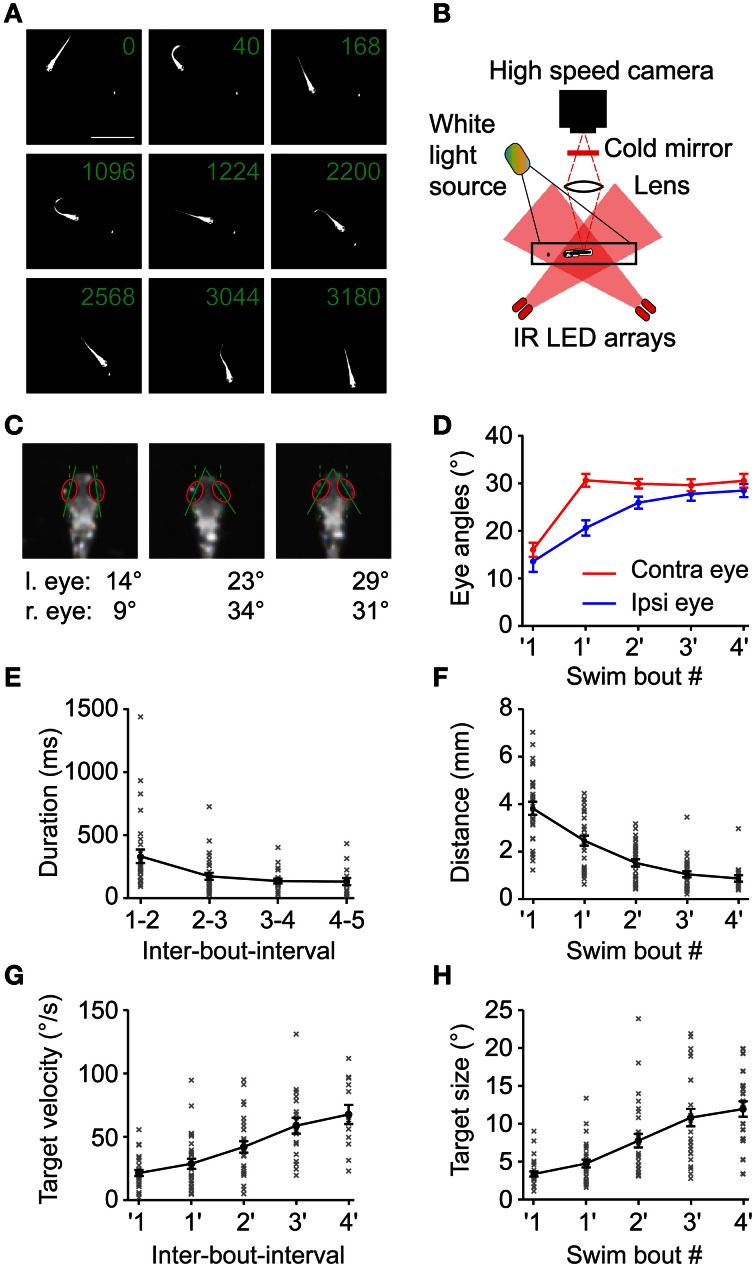
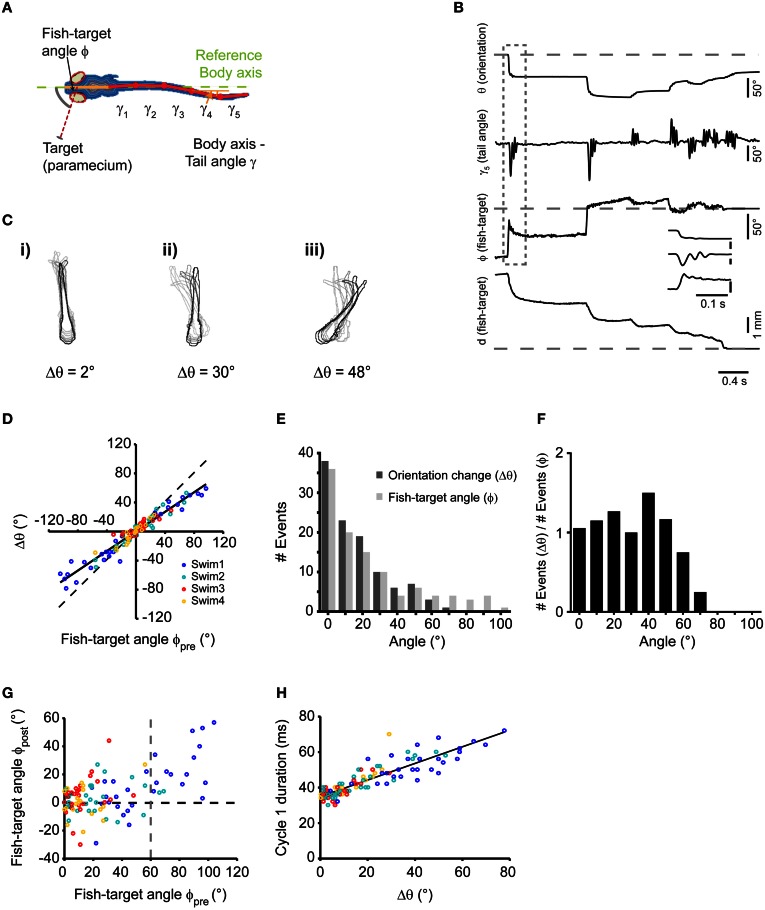
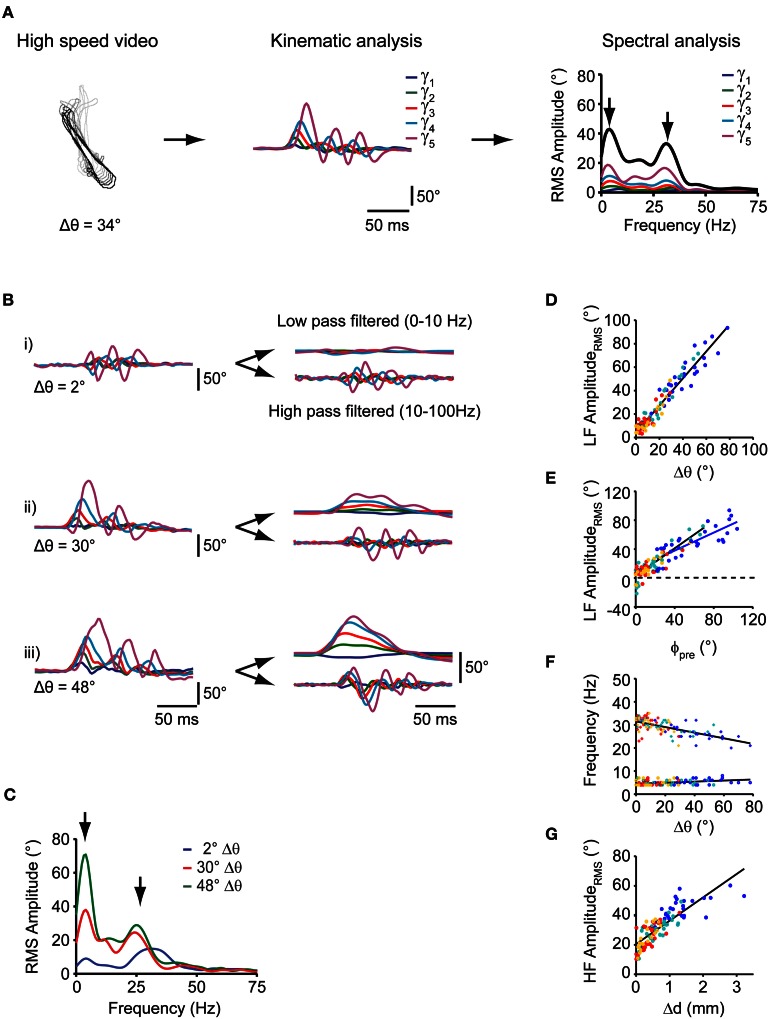
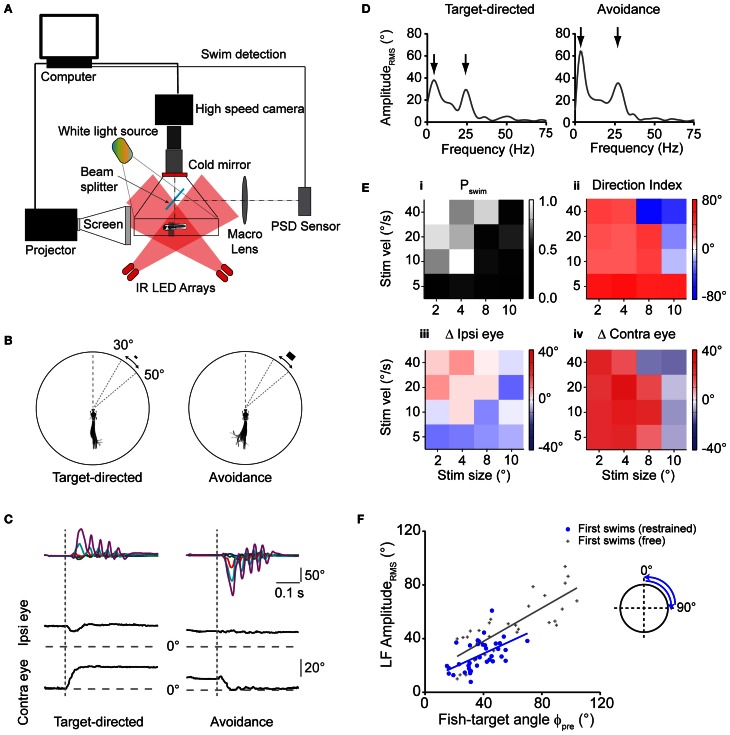
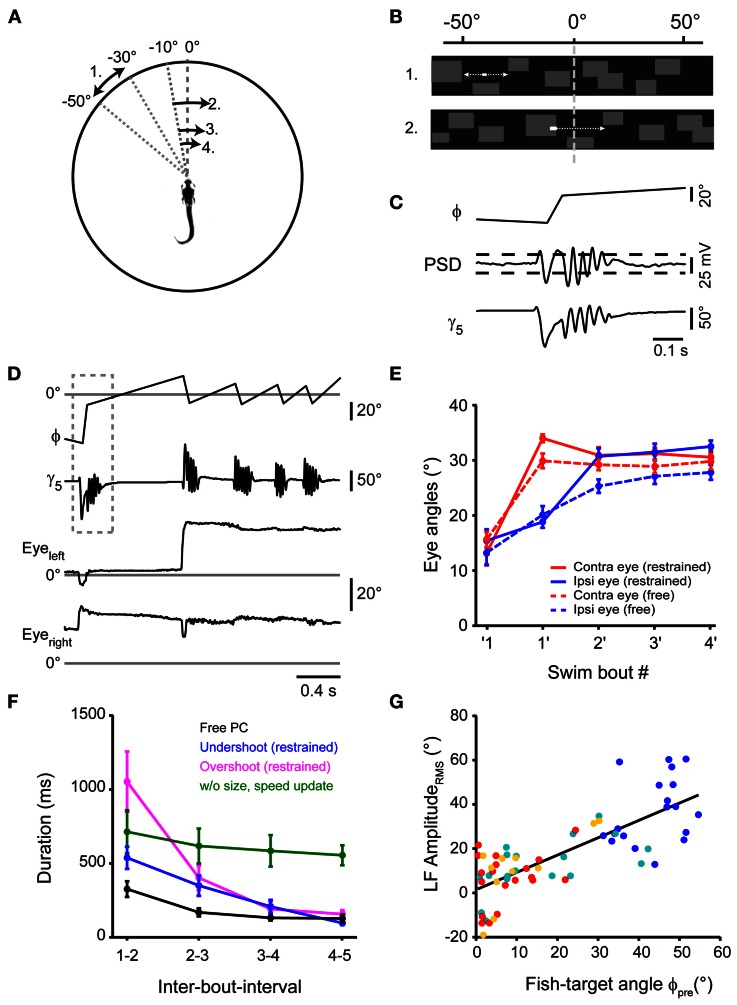
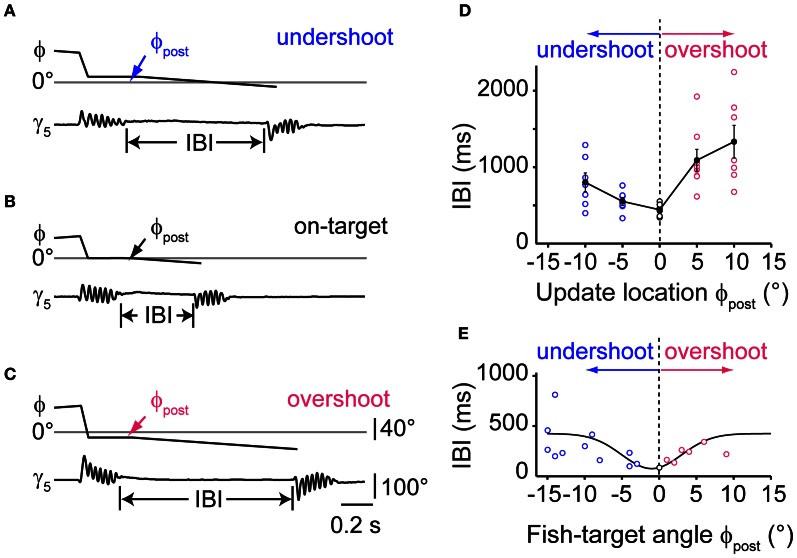
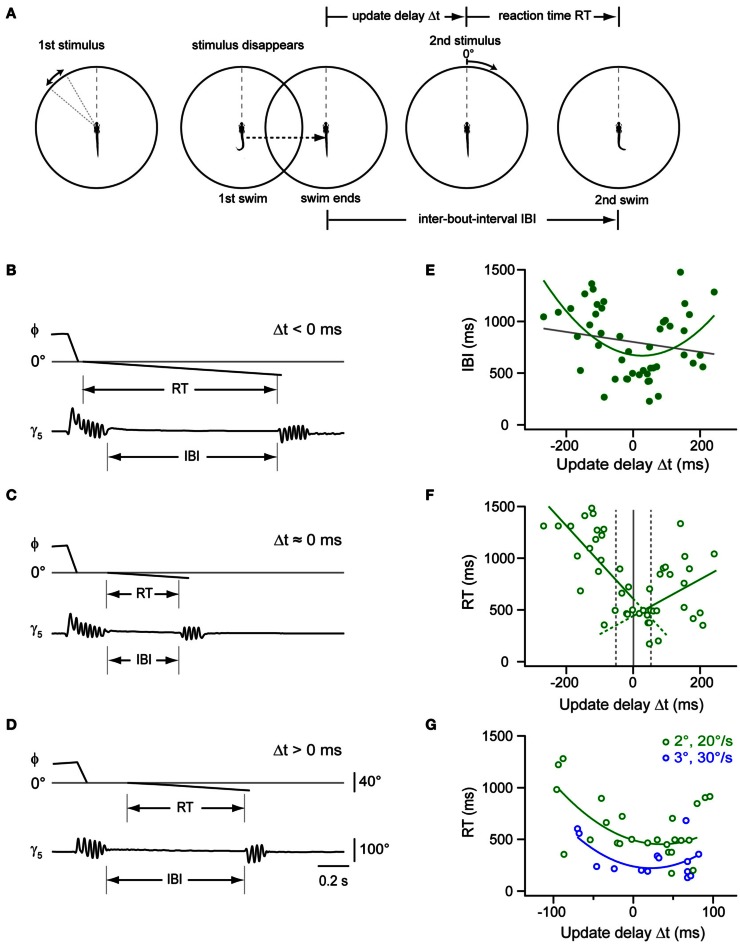
Prey capture behavior critically depends on rapid processing of sensory input in order to track, approach, and catch the target. When using vision, the nervous system faces the problem of extracting relevant information from a continuous stream of input in order to detect and categorize visible objects as potential prey and to select appropriate motor patterns for approach. For prey capture, many vertebrates exhibit intermittent locomotion, in which discrete motor patterns are chained into a sequence, interrupted by short periods of rest. Here, using high-speed recordings of full-length prey capture sequences performed by freely swimming zebrafish larvae in the presence of a single paramecium, we provide a detailed kinematic analysis of first and subsequent swim bouts during prey capture. Using Fourier analysis, we show that individual swim bouts represent an elementary motor pattern. Changes in orientation are directed toward the target on a graded scale and are implemented by an asymmetric tail bend component superimposed on this basic motor pattern. To further investigate the role of visual feedback on the efficiency and speed of this complex behavior, we developed a closed-loop virtual reality setup in which minimally restrained larvae recapitulated interconnected swim patterns closely resembling those observed during prey capture in freely moving fish. Systematic variation of stimulus properties showed that prey capture is initiated within a narrow range of stimulus size and velocity. Furthermore, variations in the delay and location of swim triggered visual feedback showed that the reaction time of secondary and later swims is shorter for stimuli that appear within a narrow spatio-temporal window following a swim. This suggests that the larva may generate an expectation of stimulus position, which enables accelerated motor sequencing if the expectation is met by appropriate visual feedback.
Keywords: double-step saccade; goal-directed behavior; intermittent locomotion; motor sequence; prey capture; saccadic suppression; virtual reality; zebrafish.
Figures
Similar articles
-
Deconstructing Hunting Behavior Reveals a Tightly Coupled Stimulus-Response Loop.Curr Biol. 2020 Jan 6;30(1):54-69.e9. doi: 10.1016/j.cub.2019.11.022. Epub 2019 Dec 19. Curr Biol. 2020. PMID: 31866365
-
Visually guided gradation of prey capture movements in larval zebrafish.J Exp Biol. 2013 Aug 15;216(Pt 16):3071-83. doi: 10.1242/jeb.087742. Epub 2013 Apr 25. J Exp Biol. 2013. PMID: 23619412 Free PMC article.
-
Prey capture by larval zebrafish: evidence for fine axial motor control.Brain Behav Evol. 2002;60(4):207-29. doi: 10.1159/000066699. Brain Behav Evol. 2002. PMID: 12457080
-
Prey capture in zebrafish larvae serves as a model to study cognitive functions.Front Neural Circuits. 2013 Jun 11;7:110. doi: 10.3389/fncir.2013.00110. eCollection 2013. Front Neural Circuits. 2013. PMID: 23781176 Free PMC article. Review.
-
[Visual system and prey capture behavior of larval zebrafish].Yi Chuan. 2013 Apr;35(4):468-76. doi: 10.3724/sp.j.1005.2013.00468. Yi Chuan. 2013. PMID: 23659937 Review. Chinese.
Cited by
-
The tectonigral pathway regulates appetitive locomotion in predatory hunting in mice.Nat Commun. 2021 Jul 20;12(1):4409. doi: 10.1038/s41467-021-24696-3. Nat Commun. 2021. PMID: 34285209 Free PMC article.
-
Pretectal neurons control hunting behaviour.Elife. 2019 Oct 8;8:e48114. doi: 10.7554/eLife.48114. Elife. 2019. PMID: 31591961 Free PMC article.
-
Visuomotor transformations underlying hunting behavior in zebrafish.Curr Biol. 2015 Mar 30;25(7):831-46. doi: 10.1016/j.cub.2015.01.042. Epub 2015 Mar 5. Curr Biol. 2015. PMID: 25754638 Free PMC article.
-
A detailed quantification of larval zebrafish behavioral repertoire uncovers principles of hunting behavior.iScience. 2025 Mar 13;28(4):112213. doi: 10.1016/j.isci.2025.112213. eCollection 2025 Apr 18. iScience. 2025. PMID: 40235586 Free PMC article.
-
From perception to behavior: The neural circuits underlying prey hunting in larval zebrafish.Front Neural Circuits. 2023 Feb 1;17:1087993. doi: 10.3389/fncir.2023.1087993. eCollection 2023. Front Neural Circuits. 2023. PMID: 36817645 Free PMC article. Review.
References
-
- Avery R., Mueller C., Smith J., Bond D. (1987). The movement patterns of lacertid lizards: speed, gait and pauses in Lacerta vivipara. J. Zool. 211, 47–63
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous