

Notch signaling maintains neural rosette polarity
- PMID: 23675446
- PMCID: PMC3651093
- DOI: 10.1371/journal.pone.0062959
Notch signaling maintains neural rosette polarity
Abstract
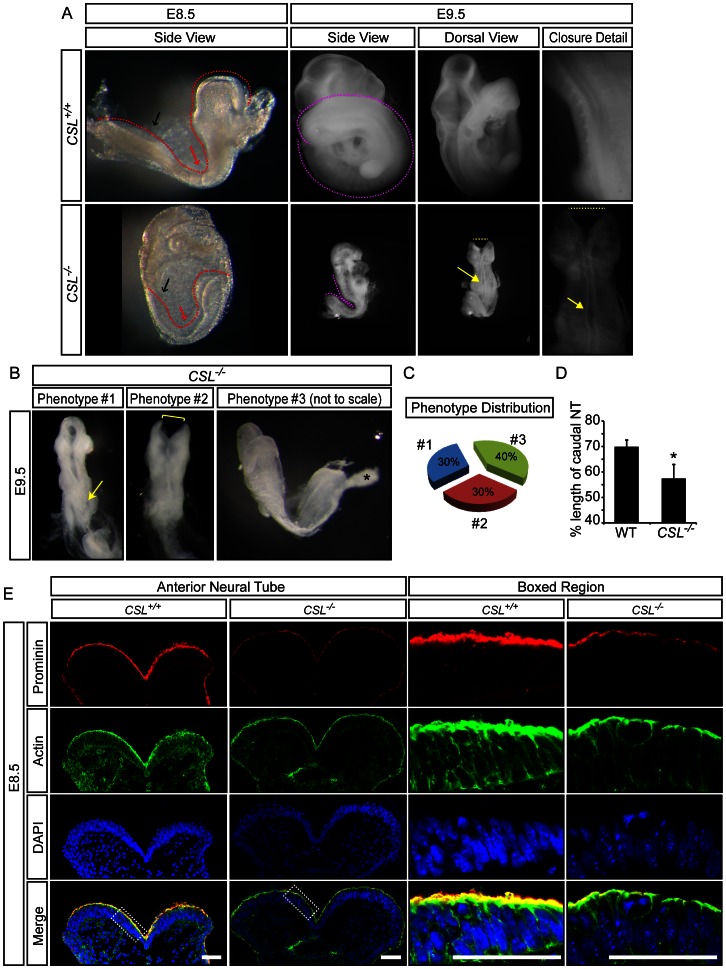
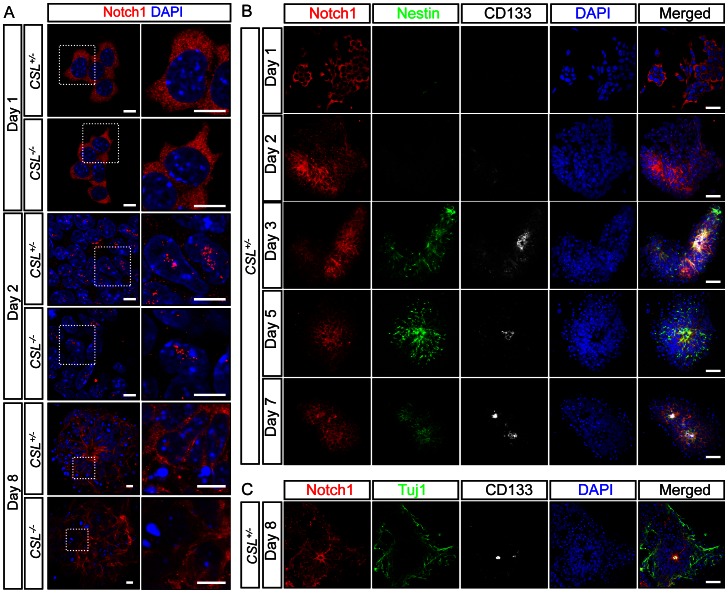
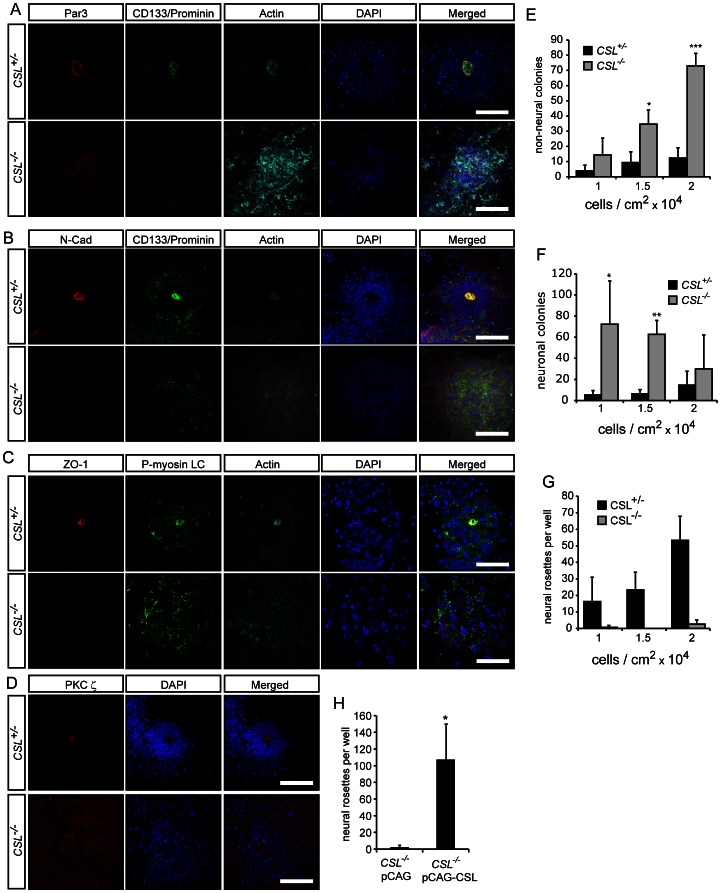
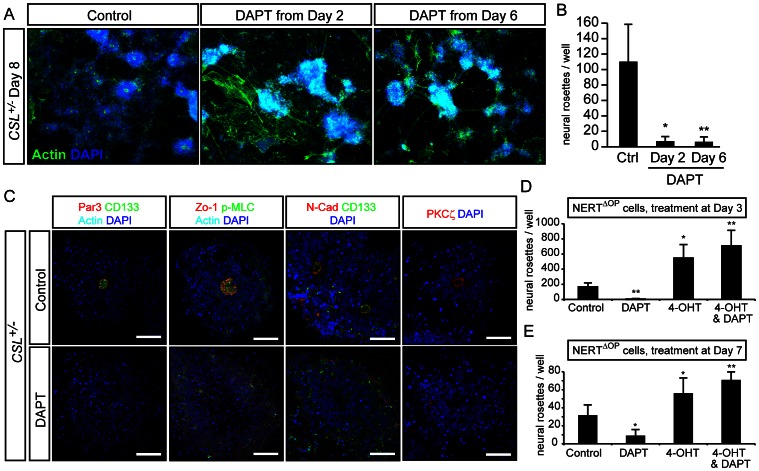
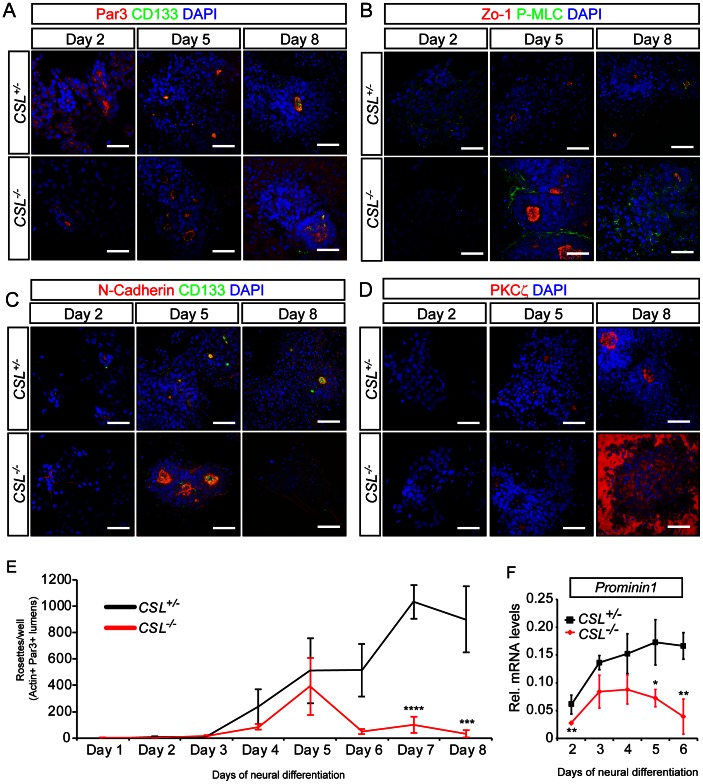
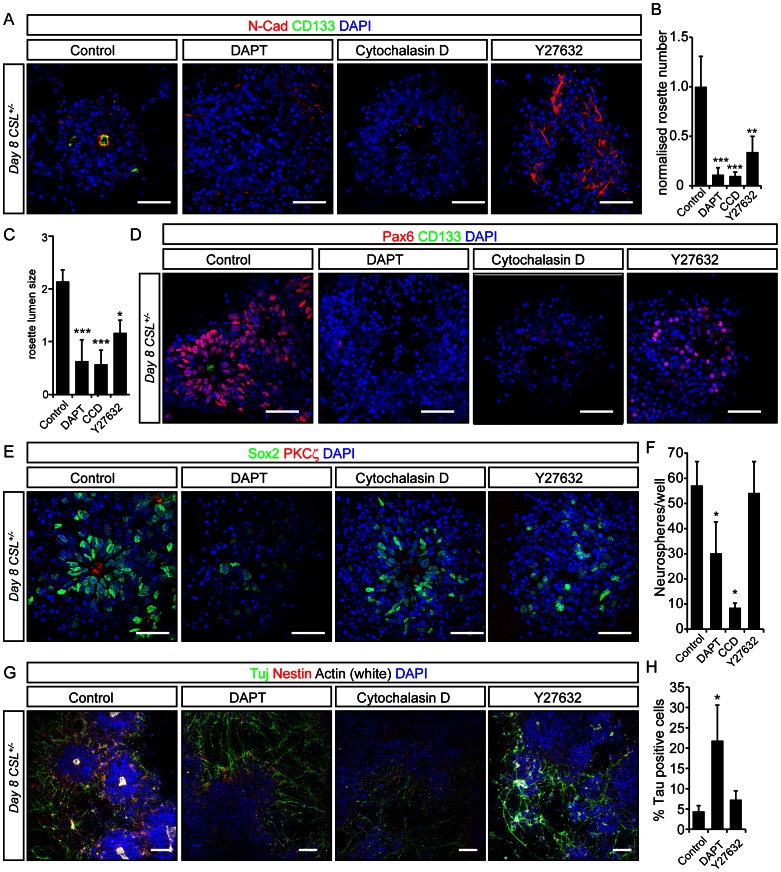
Formation of the metazoan body plan requires a complex interplay of morphological changes and patterning, and central to these processes is the establishment of apical/basal cell polarity. In the developing nervous system, apical/basal cell polarity is essential for neural tube closure and maintenance of the neural stem cell population. In this report we explore how a signaling pathway important for nervous system development, Notch signaling, impacts on apical/basal cell polarity in neural differentiation. CSL(-/-) mouse embryos, which are devoid of canonical Notch signaling, demonstrated a neural tube phenotype consistent with cell polarity and convergent extension defects, including deficiencies in the restricted expression of apical polarity markers in the neuroepithelium. CSL(-/-) mouse embryonic stem (ES) cells, cultured at low density, behaved as wild-type in the establishment of neural progenitors and apical specification, though progression through rosette formation, an in vitro correlate of neurulation, required CSL for correct maintenance of rosette structure and regulation of neuronal differentiation. Similarly, acute pharmacological inhibition of Notch signaling led to the breakdown of neural rosettes and accelerated neuronal differentiation. In addition to functional Notch signaling, rosette integrity was found to require actin polymerization and Rho kinase (ROCK) activity. Disruption of rosettes through inhibition of actin polymerization or ROCK activity, however, had no effect on neuronal differentiation, indicating that rosette maintenance is not a prerequisite for normal neuronal differentiation. In conclusion, our data indicate that Notch signaling plays a role not only in differentiation, but also in organization and maintenance of polarity during development of the early nervous system.
Conflict of interest statement
Figures

References
-
- Copp AJ, Greene ND, Murdoch JN (2003) The genetic basis of mammalian neurulation. Nat Rev Genet 4: 784–793. - PubMed
-
- Mitchell KJ (2011) The genetics of neurodevelopmental disease. Curr Opin Neurobiol 21: 197–203. - PubMed
-
- Doudney K, Stanier P (2005) Epithelial cell polarity genes are required for neural tube closure. Am J Med Genet C Semin Med Genet 135C: 42–47. - PubMed
-
- Desmond ME, Schoenwolf GC (1986) Evaluation of the roles of intrinsic and extrinsic factors in occlusion of the spinal neurocoel during rapid brain enlargement in the chick embryo. J Embryol Exp Morphol 97: 25–46. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
