

Glia maturation factor-γ negatively modulates TLR4 signaling by facilitating TLR4 endocytic trafficking in macrophages
- PMID: 23677465
- PMCID: PMC3679304
- DOI: 10.4049/jimmunol.1203048
Glia maturation factor-γ negatively modulates TLR4 signaling by facilitating TLR4 endocytic trafficking in macrophages
Abstract
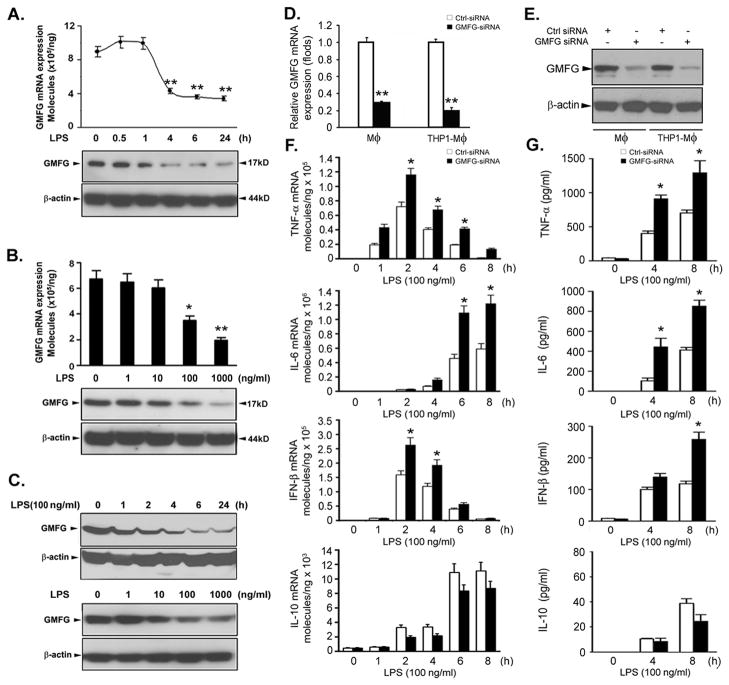
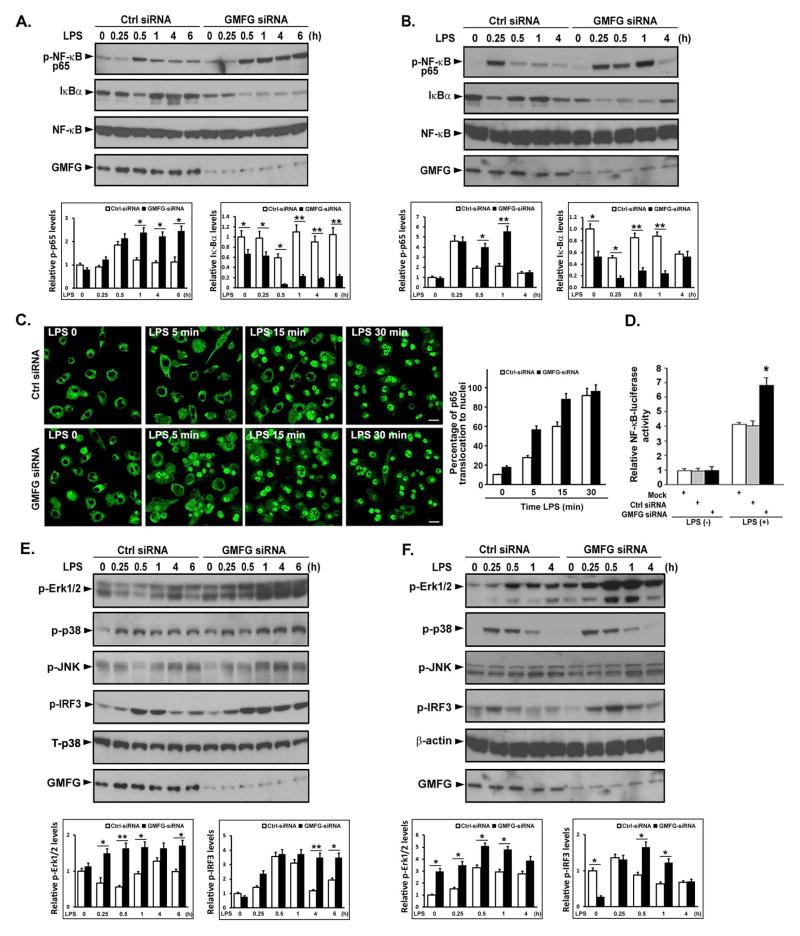
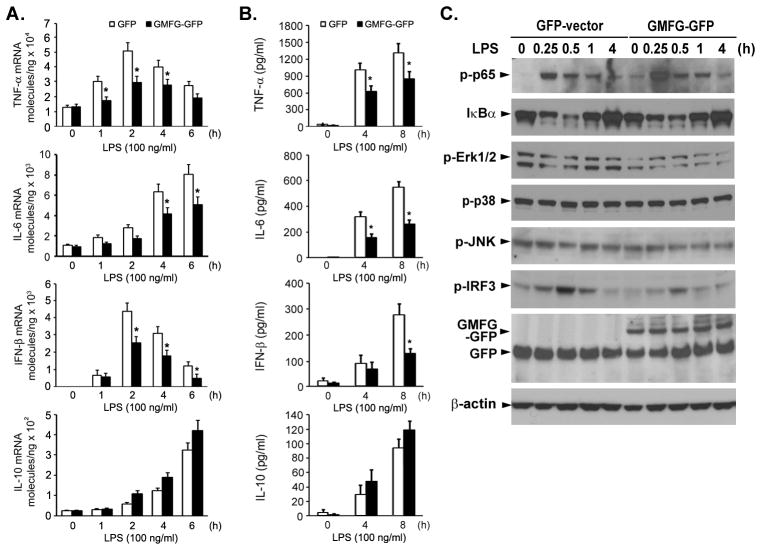
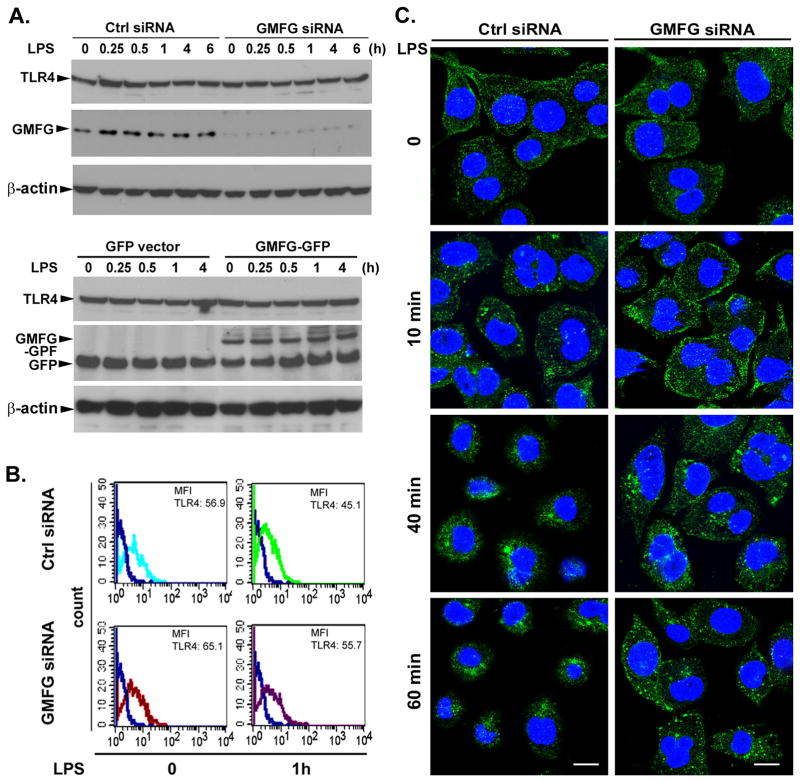
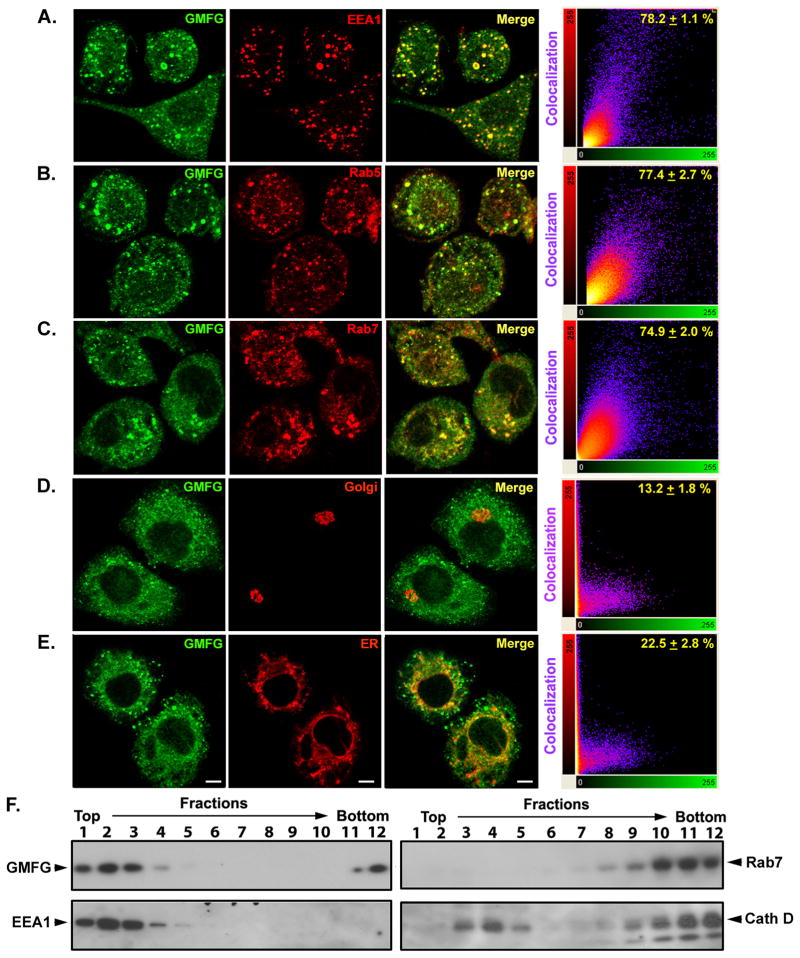
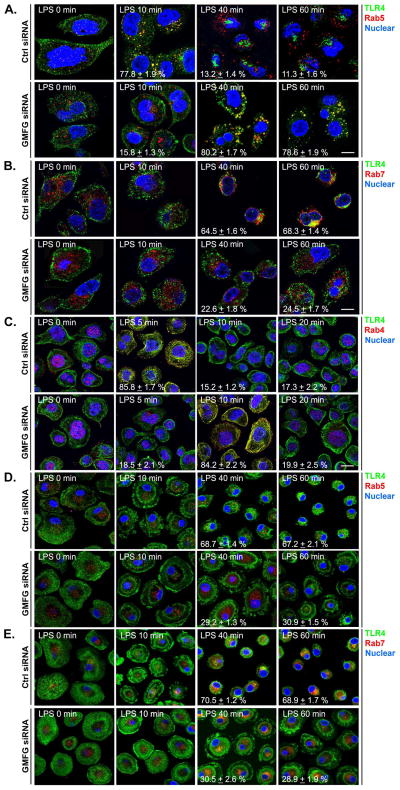
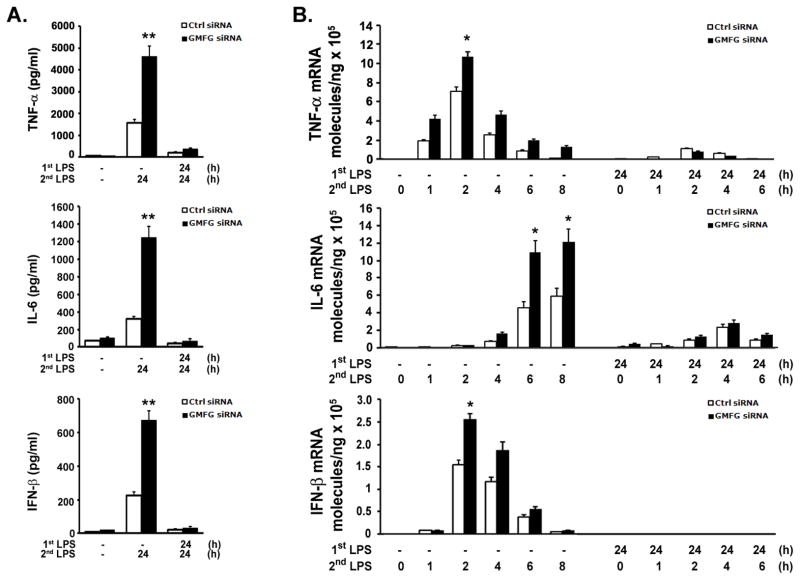
TLR4 signaling must be tightly regulated to provide both effective immune protection and avoid inflammation-induced pathology. Thus, the mechanisms that negatively regulate the TLR4-triggered inflammatory response are of particular importance. Glia maturation factor-γ (GMFG), a novel actin depolymerization factor/cofilin superfamily protein that is expressed in inflammatory cells, has been implicated in mediating neutrophil and T cell migration, but its function in macrophage immune response remains unclear. In the current study, the role of GMFG in the LPS-induced TLR4-signaling pathway was investigated in THP-1 macrophages and human primary macrophages. LPS stimulation of macrophages decreased GMFG mRNA and protein expression. We show that GMFG negatively regulates LPS-induced activation of NF-κB-, MAPK-, and IRF3-signaling pathways and subsequent production of proinflammatory cytokines and type I IFN in human macrophages. We found that endogenous GMFG localized within early and late endosomes. GMFG knockdown delayed LPS-induced TLR4 internalization and caused prolonged TLR4 retention at the early endosome, suggesting that TLR4 transport from early to late endosomes is interrupted, which may contribute to enhanced LPS-induced TLR4 signaling. Taken together, our findings suggest that GMFG functions as a negative regulator of TLR4 signaling by facilitating TLR4 endocytic trafficking in macrophages.
Conflict of interest statement
The authors have no conflicting financial interests.
Figures
References
-
- Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006;124:783–801. - PubMed
-
- West AP, Koblansky AA, Ghosh S. Recognition and signaling by toll-like receptors. Annu Rev Cell Dev Biol. 2006;22:409–437. - PubMed
-
- Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol. 2010;11:373–384. - PubMed
-
- Akira S, Takeda K. Toll-like receptor signalling. Nat Rev Immunol. 2004;4:499–511. - PubMed
-
- Barrat FJ, Coffman RL. Development of TLR inhibitors for the treatment of autoimmune diseases. Immunol Rev. 2008;223:271–283. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
