

Identification of a highly conserved surface on Tat variants
- PMID: 23678001
- PMCID: PMC3696680
- DOI: 10.1074/jbc.M113.466011
Identification of a highly conserved surface on Tat variants
Abstract
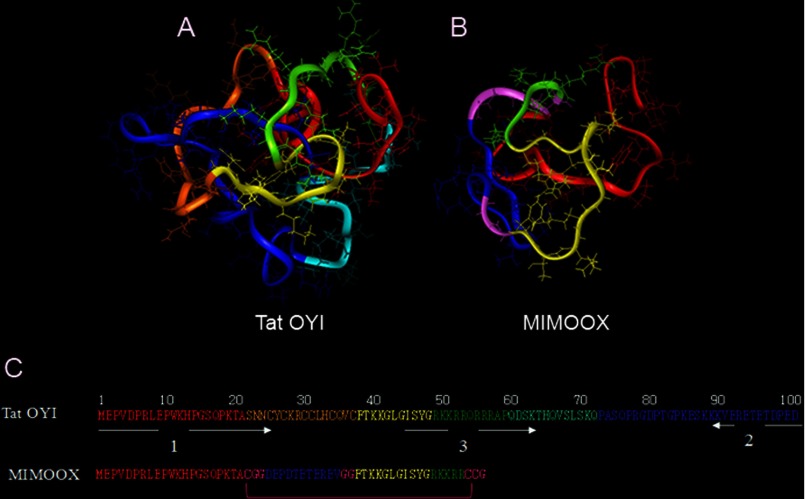
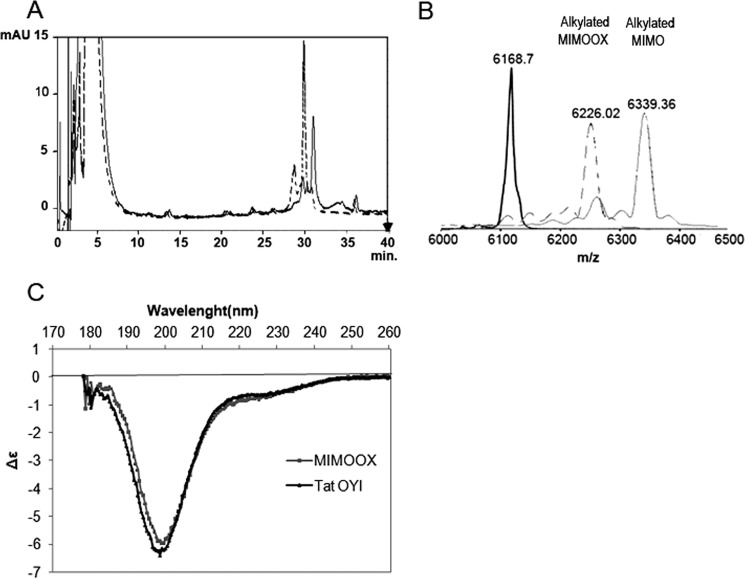
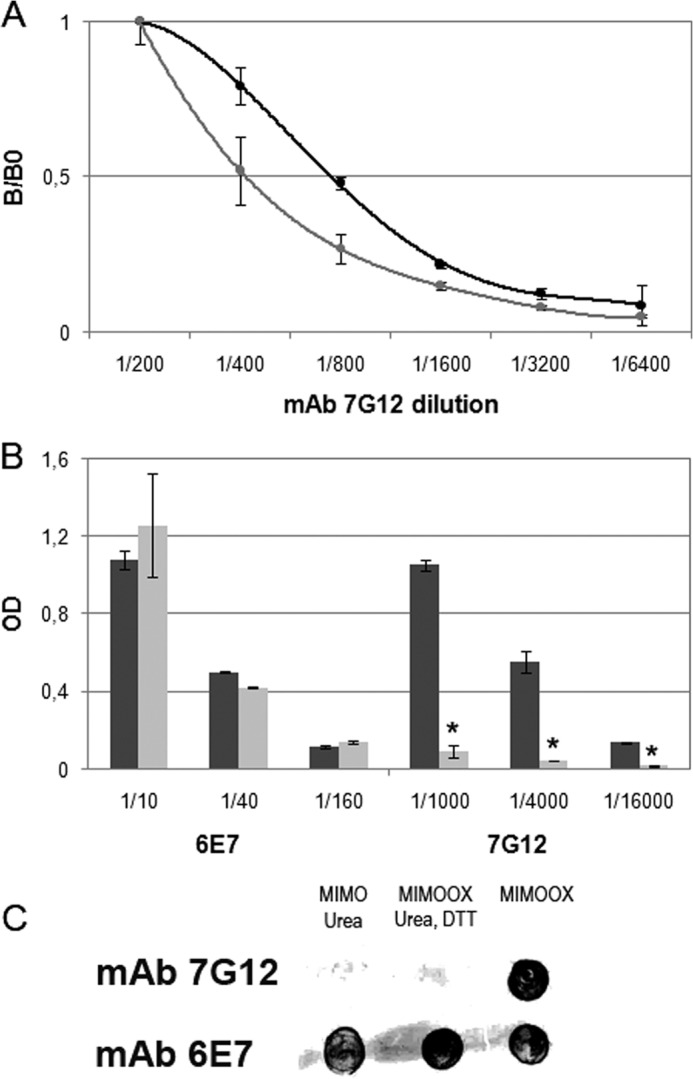
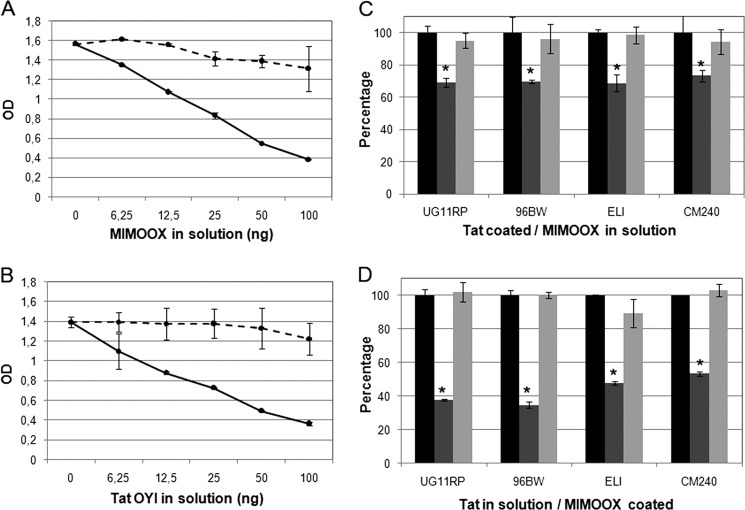
Extracellular Tat is suspected to protect HIV-1-infected cells from cellular immunity. Seropositive patients are unable to produce neutralizing antibodies against Tat, and Tat is still secreted under antiviral treatment. In mice, the Tat OYI vaccine candidate generates neutralizing antibodies such as the mAb 7G12. A peptide called MIMOOX was designed from fragments of Tat OYI identified as the possible binding site for mAb 7G12. MIMOOX was chemically synthesized, and its structure was stabilized with a disulfide bridge. Circular dichroism spectra showed that MIMOOX had mainly β turns but no α helix as Tat OYI. MIMOOX was recognized by mAb 7G12 in ELISA only in reduced conditions. Moreover, a competitive recognition assay with mAb 7G12 between MIMOOX and Tat variants showed that MIMOOX mimics a highly conserved surface in Tat variants. Rat immunizations with MIMOOX induce antibodies recognizing Tat variants from the main HIV-1 subtypes and confirm the Tat OYI vaccine approach.
Keywords: Cellular Immune Response; Epitope Mapping; HIV-1; Protein Design; Tat; Vaccine; Vaccine Development.
Figures
Similar articles
-
A monoclonal antibody directed against a conformational epitope of the HIV-1 trans-activator (Tat) protein neutralizes cross-clade.J Biol Chem. 2012 Apr 6;287(15):11942-50. doi: 10.1074/jbc.M111.319863. Epub 2012 Feb 23. J Biol Chem. 2012. PMID: 22362765 Free PMC article.
-
Fab'-induced folding of antigenic N-terminal peptides from intrinsically disordered HIV-1 Tat revealed by X-ray crystallography.J Mol Biol. 2011 Jan 7;405(1):33-42. doi: 10.1016/j.jmb.2010.10.033. Epub 2010 Oct 28. J Mol Biol. 2011. PMID: 21035463 Review.
-
Reservoir cells no longer detectable after a heterologous SHIV challenge with the synthetic HIV-1 Tat Oyi vaccine.Retrovirology. 2006 Jan 27;3:8. doi: 10.1186/1742-4690-3-8. Retrovirology. 2006. PMID: 16441880 Free PMC article.
-
Tat HIV-1 primary and tertiary structures critical to immune response against non-homologous variants.J Biol Chem. 2002 Sep 27;277(39):35915-9. doi: 10.1074/jbc.M204393200. Epub 2002 Jun 21. J Biol Chem. 2002. PMID: 12080071
-
What does the structure-function relationship of the HIV-1 Tat protein teach us about developing an AIDS vaccine?Retrovirology. 2009 May 25;6:50. doi: 10.1186/1742-4690-6-50. Retrovirology. 2009. PMID: 19467159 Free PMC article. Review.
Cited by
-
Multimodality vaccination against clade C SHIV: partial protection against mucosal challenges with a heterologous tier 2 virus.Vaccine. 2014 Nov 12;32(48):6527-36. doi: 10.1016/j.vaccine.2014.08.065. Epub 2014 Sep 20. Vaccine. 2014. PMID: 25245933 Free PMC article.
-
HIV Tat protein and amyloid-β peptide form multifibrillar structures that cause neurotoxicity.Nat Struct Mol Biol. 2017 Apr;24(4):379-386. doi: 10.1038/nsmb.3379. Epub 2017 Feb 20. Nat Struct Mol Biol. 2017. PMID: 28218748 Free PMC article.
-
Intradermal injection of a Tat Oyi-based therapeutic HIV vaccine reduces of 1.5 log copies/mL the HIV RNA rebound median and no HIV DNA rebound following cART interruption in a phase I/II randomized controlled clinical trial.Retrovirology. 2016 Apr 1;13:21. doi: 10.1186/s12977-016-0251-3. Retrovirology. 2016. PMID: 27036656 Free PMC article. Clinical Trial.
-
Didehydro-Cortistatin A Inhibits HIV-1 by Specifically Binding to the Unstructured Basic Region of Tat.mBio. 2019 Feb 5;10(1):e02662-18. doi: 10.1128/mBio.02662-18. mBio. 2019. PMID: 30723126 Free PMC article.
References
-
- Wong-Staal F., Gallo R. C. (1985) Human T-lymphotropic retroviruses. Nature 317, 395–403 - PubMed
-
- Debaisieux S., Rayne F., Yezid H., Beaumelle B. (2012) The ins and outs of HIV-1 Tat. Traffic 13, 355–363 - PubMed
-
- Westendorp M. O., Frank R., Ochsenbauer C., Stricker K., Dhein J., Walczak H., Debatin K. M., Krammer P. H. (1995) Sensitization of T cells to CD95-mediated apoptosis by HIV-1 Tat and gp120. Nature 375, 497–500 - PubMed
-
- Mediouni S., Darque A., Baillat G., Ravaux I., Dhiver C., Tissot-Dupont H., Mokhtari M., Moreau H., Tamalet C., Brunet C., Paul P., Dignat-George F., Stein A., Brouqui P., Spector S. A., Campbell G. R., Loret E. P. (2012a) Antiretroviral therapy does not block the secretion of the human immunodeficiency virus tat protein. Infect. Disord. Drug Targets 12, 81–86 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
