

Superoxide poisons mononuclear iron enzymes by causing mismetallation
- PMID: 23678969
- PMCID: PMC3731988
- DOI: 10.1111/mmi.12263
Superoxide poisons mononuclear iron enzymes by causing mismetallation
Abstract
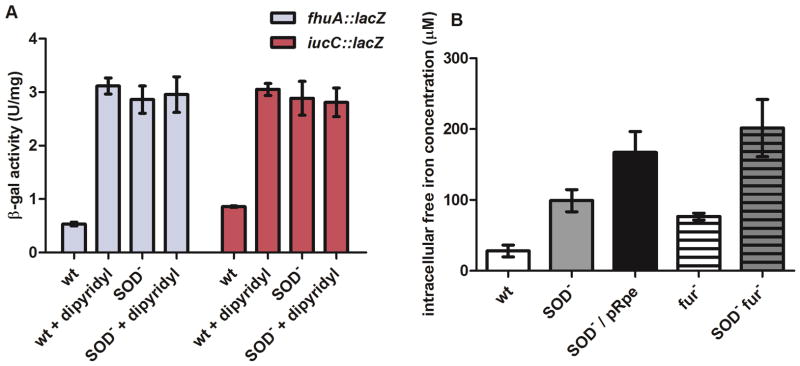
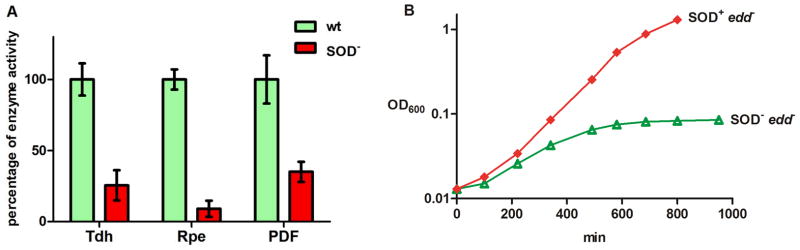
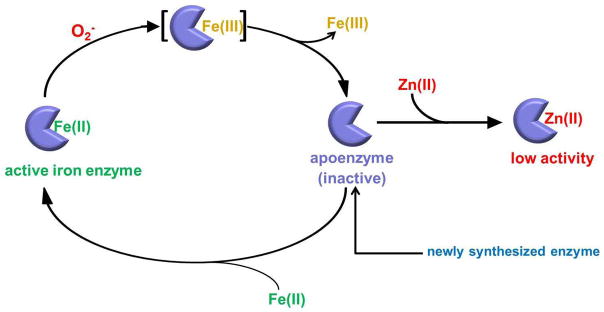
Superoxide (O(2)(-)) is a primary agent of intracellular oxidative stress. Genetic studies in many organisms have confirmed that excess O(2)(-) disrupts metabolism, but to date only a small family of [4Fe-4S] dehydratases have been identified as direct targets. This investigation reveals that in Escherichia coli O(2)(-) also poisons a broader cohort of non-redox enzymes that employ ferrous iron atoms as catalytic cofactors. These enzymes were inactivated by O(2)(-) both in vitro and in vivo. Although the enzymes are known targets of hydrogen peroxide, the outcome with O(2)(-) differs substantially. When purified enzymes were damaged by O(2)(-) in vitro, activity could be completely restored by iron addition, indicating that the O(2)(-) treatment generated an apoprotein without damaging the protein polypeptide. Superoxide stress inside cells caused the progressive mismetallation of these enzymes with zinc, which confers little activity. When O(2)(-) stress was terminated, cells gradually restored activity by extracting zinc from the proteins. The overloading of cells with zinc caused mismetallation even without O(2)(-) stress. These results support a model in which O(2)(-) repeatedly excises iron from these enzymes, allowing zinc to compete with iron for remetallation of their apoprotein forms. This action substantially expands the physiological imprint of O(2)(-) stress.
© 2013 John Wiley & Sons Ltd.
Figures





References
-
- Akana J, Fedorov AA, Fedorov E, Novak WR, Babbitt PC, Almo SC, Gerlt JA. D-ribulose 5-phosphate 3-epimerase: functional and structural relationships to members of the ribulose-phosphate binding (β/α) 8-barrel superfamily. Biochemistry. 2006;45:2493–2503. - PubMed
-
- Al-Maghrebi M, Fridovich I, Benov L. Manganese supplementation relieves the phenotypic deficits seen in superoxide-dismutase-null Escherichia coli. Arch Biochem Biophys. 2002;402:104–109. - PubMed
-
- Altuvia S, Almiron M, Huisman G, Kolter R, Storz G. The dps promoter is activated by OxyR during growth and by IHF and sigma S in stationary phase. Mol Microbiol. 1994;13:265–272. - PubMed
-
- Anbar AD. Elements and evolution. Science. 2008;322:1481–1483. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
