

Plant defense against insect herbivores
- PMID: 23681010
- PMCID: PMC3676838
- DOI: 10.3390/ijms140510242
Plant defense against insect herbivores
Abstract
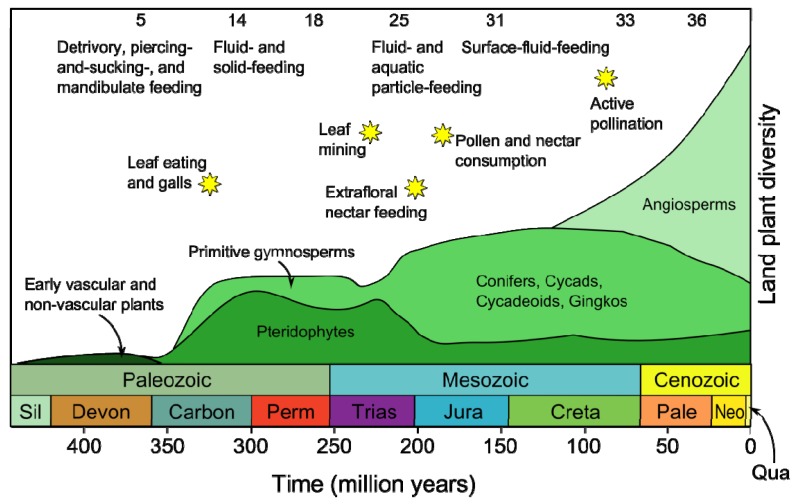
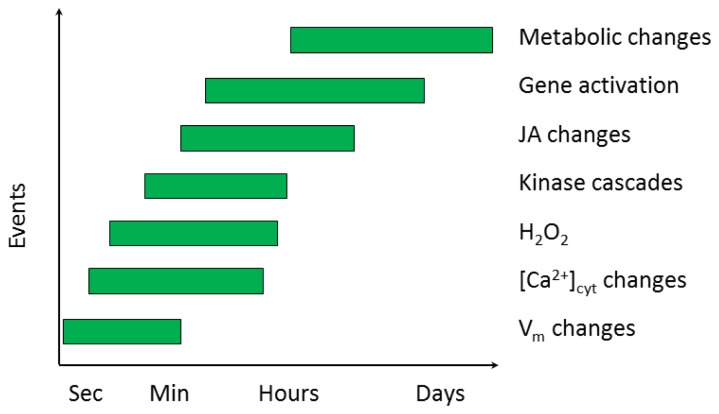
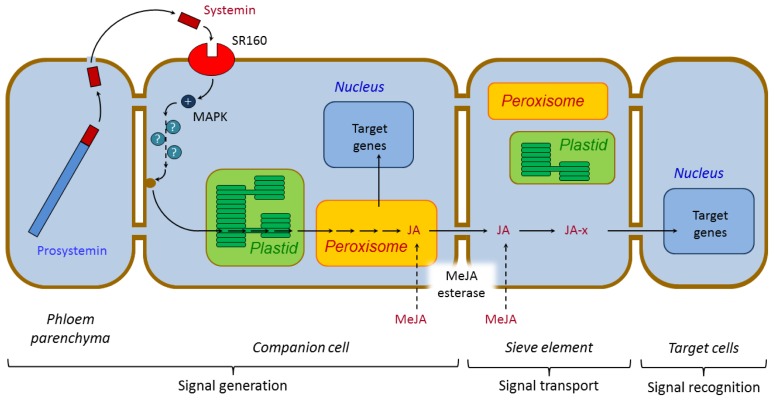
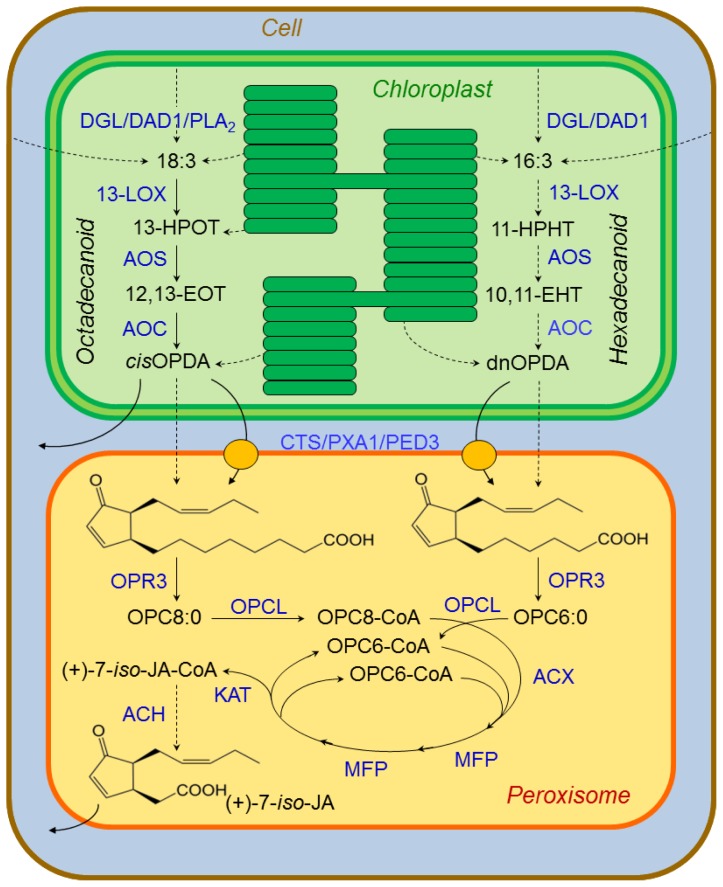
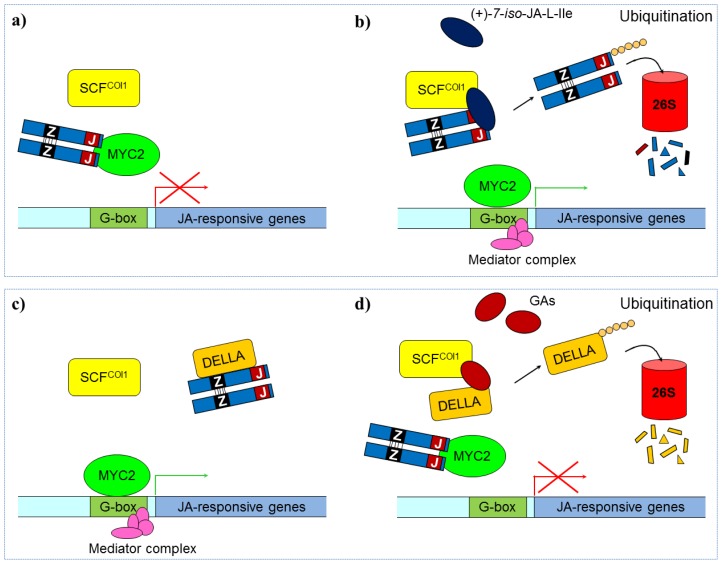
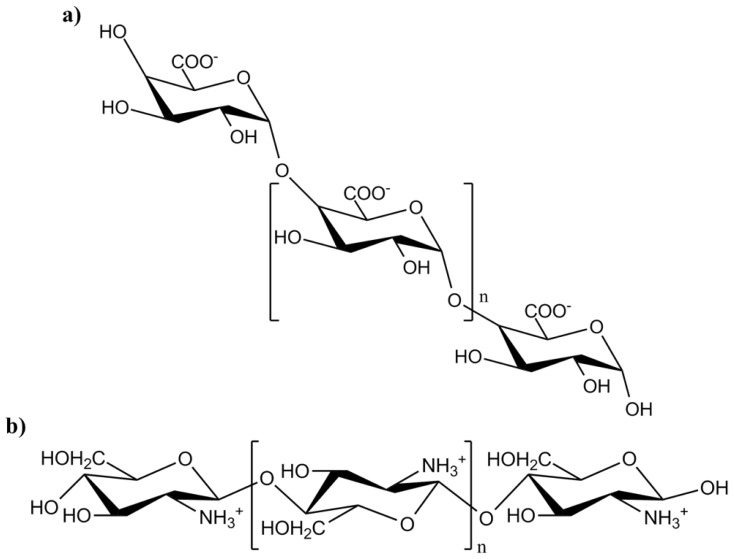
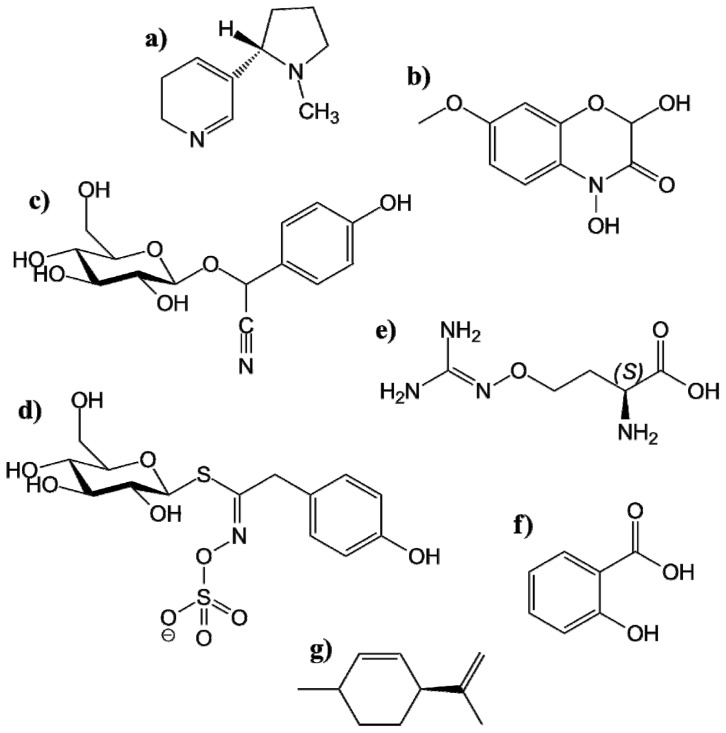
Plants have been interacting with insects for several hundred million years, leading to complex defense approaches against various insect feeding strategies. Some defenses are constitutive while others are induced, although the insecticidal defense compound or protein classes are often similar. Insect herbivory induce several internal signals from the wounded tissues, including calcium ion fluxes, phosphorylation cascades and systemic- and jasmonate signaling. These are perceived in undamaged tissues, which thereafter reinforce their defense by producing different, mostly low molecular weight, defense compounds. These bioactive specialized plant defense compounds may repel or intoxicate insects, while defense proteins often interfere with their digestion. Volatiles are released upon herbivory to repel herbivores, attract predators or for communication between leaves or plants, and to induce defense responses. Plants also apply morphological features like waxes, trichomes and latices to make the feeding more difficult for the insects. Extrafloral nectar, food bodies and nesting or refuge sites are produced to accommodate and feed the predators of the herbivores. Meanwhile, herbivorous insects have adapted to resist plant defenses, and in some cases even sequester the compounds and reuse them in their own defense. Both plant defense and insect adaptation involve metabolic costs, so most plant-insect interactions reach a stand-off, where both host and herbivore survive although their development is suboptimal.
Figures
References
-
- Ehrlich P.R., Raven P.H. Butterflies and plants: A study in coevolution. Evolution. 1964;18:586–608.
-
- Labandeira C.C. Insect mouthparts: Ascertaining the paleobiology of insect feeding strategies. Annu. Rev. Ecol. Syst. 1997;28:153–193.
-
- Niklas K.J., Tiffney B.H., Knoll A.H. Patterns in Vascular Land Plant Diversification: An Analysis at the Species Level. In: Valentine J.W., editor. Phanerozoic Diversity Patterns: Profiles in Macroevolution. Princeton University Press; Princeton, NJ, USA: 1985. pp. 97–128.
-
- Schönenberger J., Friis E.M., Matthews M.L., Endress P.K. Cunoniaceae in the cretaceous of europe: Evidence from fossil flowers. Ann. Bot. 2001;88:423–437.
-
- Schaller A., Howe G.A. Direct Defenses in Plants and Their Induction by Wounding and Insect Herbivores. In: Schaller A., editor. Induced Plant Resistance to Herbivory. Springer Science+Business Media; Berlin, Germany: 2008. pp. 7–29.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
