

Unbiased segregation of fission yeast chromosome 2 strands to daughter cells
- PMID: 23681661
- PMCID: PMC6959520
- DOI: 10.1007/s10577-013-9352-1
Unbiased segregation of fission yeast chromosome 2 strands to daughter cells
Abstract
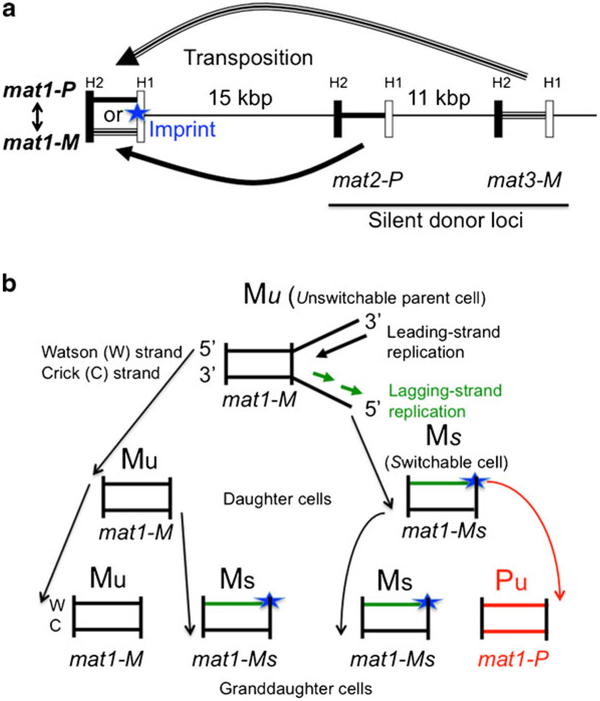
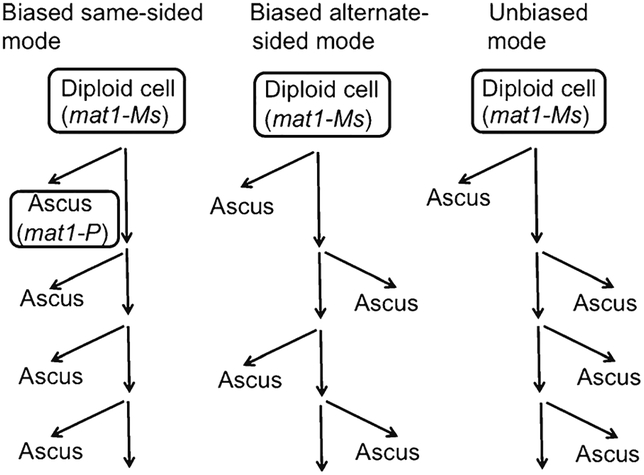
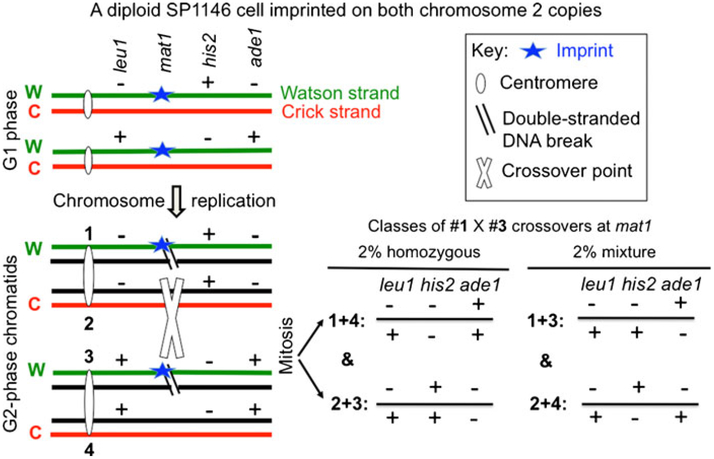
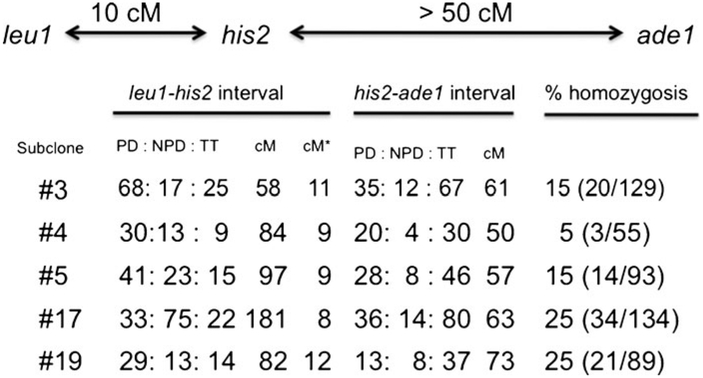
The base complementarity feature (Watson and Crick in Nature 171(4356):737-738, 1953) and the rule of semi-conservative mode of DNA replication (Messelson and Stahl in Proc Natl Acad Sci U S A 44:671-682, 1958) dictate that two identical replicas of the parental chromosome are produced during replication. In principle, the inherent strand sequence differences could generate nonequivalent daughter chromosome replicas if one of the two strands were epigenetically imprinted during replication to effect silencing/expression of developmentally important genes. Indeed, inheritance of such a strand- and site-specific imprint confers developmental asymmetry to fission yeast sister cells by a phenomenon called mating/cell-type switching. Curiously, location of DNA strands with respect to each other at the centromere is fixed, and as a result, their selected segregation to specific sister chromatid copies occurs in eukaryotic cells. The yeast system provides a unique opportunity to determine the significance of such biased strand distribution to sister chromatids. We determined whether the cylindrical-shaped yeast cell distributes the specific chromosomal strand to the same cellular pole in successive cycles of cell division. By observing the pattern of recurrent mating-type switching in progenies of individual cells by microscopic analyses, we found that chromosome 2 strands are distributed by the random mode in successive cell divisions. We also exploited unusual "hotspot" recombination features of this system to investigate whether there is selective segregation of strands such that oldest Watson-containing strands co-segregate in the diploid cell at mitosis. Our data suggests that chromosome 2 strands are segregated independently to those of the homologous chromosome.
Conflict of interest statement
Figures

Similar articles
-
Chiasmata promote monopolar attachment of sister chromatids and their co-segregation toward the proper pole during meiosis I.PLoS Genet. 2011 Mar;7(3):e1001329. doi: 10.1371/journal.pgen.1001329. Epub 2011 Mar 10. PLoS Genet. 2011. PMID: 21423721 Free PMC article.
-
Cell type regulates selective segregation of mouse chromosome 7 DNA strands in mitosis.Science. 2006 Feb 24;311(5764):1146-9. doi: 10.1126/science.1120519. Science. 2006. PMID: 16497932
-
Discovery of the mitotic selective chromatid segregation phenomenon and its implications for vertebrate development.Curr Opin Cell Biol. 2010 Feb;22(1):81-7. doi: 10.1016/j.ceb.2009.11.006. Curr Opin Cell Biol. 2010. PMID: 20022232 Free PMC article. Review.
-
Dynein participates in chromosome segregation in fission yeast.Biol Cell. 2007 Nov;99(11):627-37. doi: 10.1042/BC20070047. Biol Cell. 2007. PMID: 17561805
-
Selective chromatid segregation mechanism proposed for the human split hand/foot malformation development by chromosome 2 translocations: A perspective.Dev Biol. 2015 Dec 1;408(1):7-13. doi: 10.1016/j.ydbio.2015.10.013. Epub 2015 Oct 20. Dev Biol. 2015. PMID: 26477560 Review.
Cited by
-
Cosegregation of asymmetric features during cell division.Open Biol. 2021 Aug;11(8):210116. doi: 10.1098/rsob.210116. Epub 2021 Aug 4. Open Biol. 2021. PMID: 34343465 Free PMC article.
-
A Unique DNA Recombination Mechanism of the Mating/Cell-type Switching of Fission Yeasts: a Review.Microbiol Spectr. 2014 Oct;2(5):10.1128/microbiolspec.MDNA3-0003-2014. doi: 10.1128/microbiolspec.MDNA3-0003-2014. Microbiol Spectr. 2014. PMID: 26104357 Free PMC article. Review.
-
Asymmetric Centromeres Differentially Coordinate with Mitotic Machinery to Ensure Biased Sister Chromatid Segregation in Germline Stem Cells.Cell Stem Cell. 2019 Nov 7;25(5):666-681.e5. doi: 10.1016/j.stem.2019.08.014. Epub 2019 Sep 26. Cell Stem Cell. 2019. PMID: 31564548 Free PMC article.
References
-
- Angehrn P, Gutz H (1968) Influence of the mating type on mitotic crossing-over in Schizosaccharomyces pombe. Genetics 60:158 Abstract
-
- Armakolas A, Klar AJS (2006) Cell type regulates selective segregation of mouse chromosome 7 DNA strands in mitosis. Science 311:1146–1149 - PubMed
-
- Armakolas A, Klar AJS (2007) Left–right dynein motor implicated in selective chromatid segregation in mouse cells. Science 315:100–101 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
