

Transcriptional repression of BODENLOS by HD-ZIP transcription factor HB5 in Arabidopsis thaliana
- PMID: 23682118
- PMCID: PMC3697942
- DOI: 10.1093/jxb/ert137
Transcriptional repression of BODENLOS by HD-ZIP transcription factor HB5 in Arabidopsis thaliana
Abstract
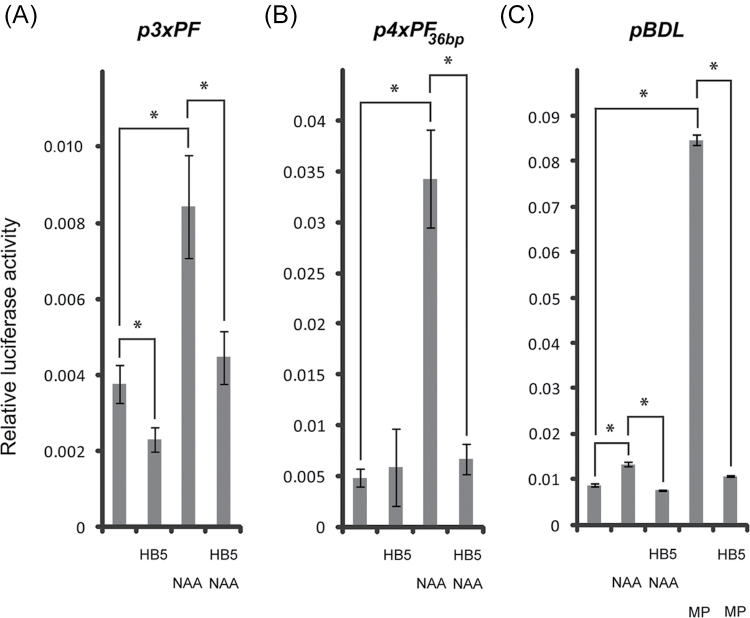
In Arabidopsis thaliana, the phytohormone auxin is an important patterning agent during embryogenesis and post-embryonic development, exerting effects through transcriptional regulation. The main determinants of the transcriptional auxin response machinery are AUXIN RESPONSE FACTOR (ARF) transcription factors and AUXIN/INDOLE-3-ACETIC ACID (AUX/IAA) inhibitors. Although members of these two protein families are major developmental regulators, the transcriptional regulation of the genes encoding them has not been well explored. For example, apart from auxin-linked regulatory inputs, factors regulating the expression of the AUX/IAA BODENLOS (BDL)/IAA12 are not known. Here, it was shown that the HOMEODOMAIN-LEUCINE ZIPPER (HD-ZIP) transcription factor HOMEOBOX PROTEIN 5 (HB5) negatively regulates BDL expression, which may contribute to the spatial control of BDL expression. As such, HB5 and probably other class I HD-ZIP proteins, appear to modulate BDL-dependent auxin response.
Keywords: Arabidopsis; BODENLOS (BDL); HOMEOBOX PROTEIN 5 (HB5); auxin; embryo; transcriptional regulation..
Figures


Similar articles
-
Auxin responsiveness of the MONOPTEROS-BODENLOS module in primary root initiation critically depends on the nuclear import kinetics of the Aux/IAA inhibitor BODENLOS.Plant J. 2016 Jan;85(2):269-77. doi: 10.1111/tpj.13108. Plant J. 2016. PMID: 26714008
-
TOPLESS mediates auxin-dependent transcriptional repression during Arabidopsis embryogenesis.Science. 2008 Mar 7;319(5868):1384-6. doi: 10.1126/science.1151461. Epub 2008 Feb 7. Science. 2008. PMID: 18258861
-
Interplay of HD-Zip II and III transcription factors in auxin-regulated plant development.J Exp Bot. 2015 Aug;66(16):5043-53. doi: 10.1093/jxb/erv174. Epub 2015 Apr 23. J Exp Bot. 2015. PMID: 25911742 Review.
-
KANADI and class III HD-Zip gene families regulate embryo patterning and modulate auxin flow during embryogenesis in Arabidopsis.Plant Cell. 2007 Feb;19(2):495-508. doi: 10.1105/tpc.106.047472. Epub 2007 Feb 16. Plant Cell. 2007. PMID: 17307928 Free PMC article.
-
Structure and function of homodomain-leucine zipper (HD-Zip) proteins.Plant Signal Behav. 2009 Feb;4(2):86-8. doi: 10.4161/psb.4.2.7692. Plant Signal Behav. 2009. PMID: 19649178 Free PMC article. Review.
Cited by
-
Molecular interactions of the γ-clade homeodomain-leucine zipper class I transcription factors during the wheat response to water deficit.Plant Mol Biol. 2016 Mar;90(4-5):435-52. doi: 10.1007/s11103-015-0427-6. Epub 2016 Jan 23. Plant Mol Biol. 2016. PMID: 26803501
-
Evolution and expression analysis reveal the potential role of the HD-Zip gene family in regulation of embryo abortion in grapes (Vitis vinifera L.).BMC Genomics. 2017 Sep 21;18(1):744. doi: 10.1186/s12864-017-4110-y. BMC Genomics. 2017. PMID: 28934927 Free PMC article.
-
Genomic survey and expression analysis of LcARFs reveal multiple functions to somatic embryogenesis in Liriodendron.BMC Plant Biol. 2024 Feb 7;24(1):94. doi: 10.1186/s12870-024-04765-7. BMC Plant Biol. 2024. PMID: 38326748 Free PMC article.
-
Genome-Wide Identification of Homeodomain Leucine Zipper (HD-ZIP) Transcription Factor, Expression Analysis, and Protein Interaction of HD-ZIP IV in Oil Palm Somatic Embryogenesis.Int J Mol Sci. 2023 Mar 5;24(5):5000. doi: 10.3390/ijms24055000. Int J Mol Sci. 2023. PMID: 36902431 Free PMC article.
-
INDEL variation in the regulatory region of the major flowering time gene LanFTc1 is associated with vernalization response and flowering time in narrow-leafed lupin (Lupinus angustifolius L.).Plant Cell Environ. 2019 Jan;42(1):174-187. doi: 10.1111/pce.13320. Epub 2018 May 23. Plant Cell Environ. 2019. PMID: 29677403 Free PMC article.
References
-
- Abel S, Nguyen MD, Theologis A. 1995. The PS-IAA4/5-like family of early auxin-inducible mRNAs in Arabidopsis thaliana . Journal of Molecular Biology 251, 533–549. - PubMed
-
- Ariel FD, Manavella PA, Dezar CA, Chan RL. 2007. The true story of the HD-Zip family. Trends in Plant Science 12, 419–426. - PubMed
-
- Baroux C, Blanvillain R, Moore IR, Gallois P. 2001. Transactivation of BARNASE under the AtLTP1 promoter affects the basal pole of the embryo and shoot development of the adult plant in Arabidopsis . The Plant Journal 28, 503–515. - PubMed
-
- Benková E, Michniewicz M, Sauer M, Teichmann T, Seifertová D, Jürgens G, Friml J. 2003. Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell 115, 591–602. - PubMed
-
- Berleth T, Jürgens G. 1993. The role of the monopteros gene in organising the basal body region of the Arabidopsis embryo. Development 118, 575–587.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
