

Regulation of the osterix (Osx, Sp7) promoter by osterix and its inhibition by parathyroid hormone
- PMID: 23682129
- PMCID: PMC3685218
- DOI: 10.1530/JME-12-0251
Regulation of the osterix (Osx, Sp7) promoter by osterix and its inhibition by parathyroid hormone
Abstract
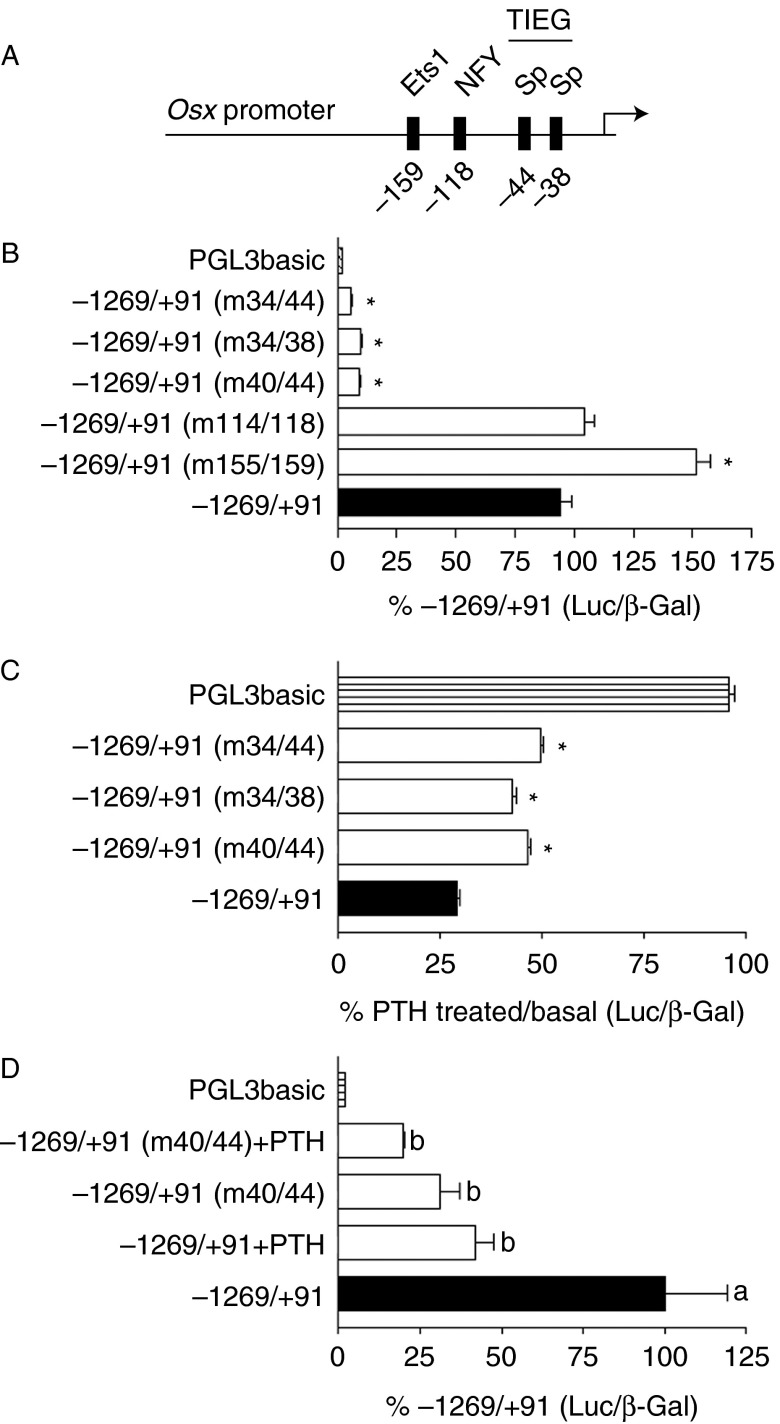
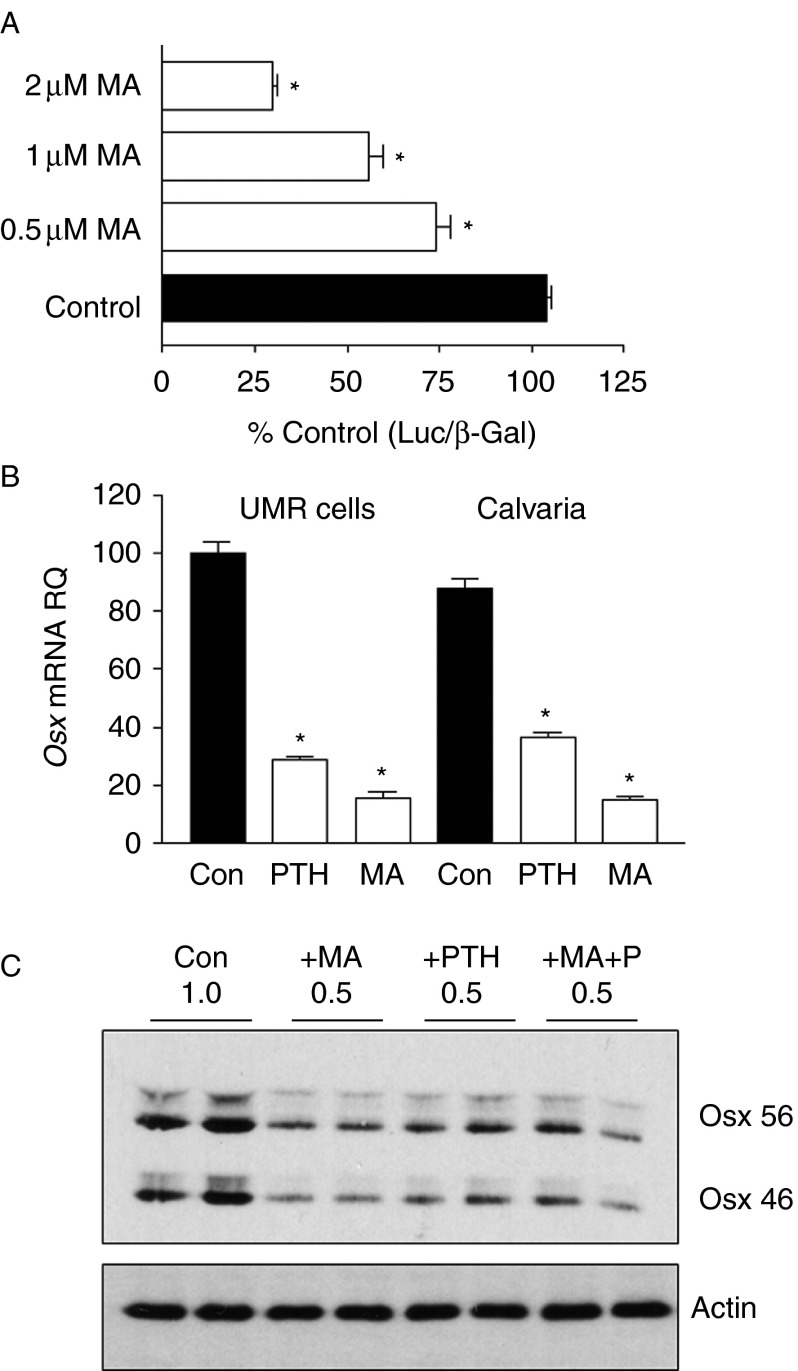
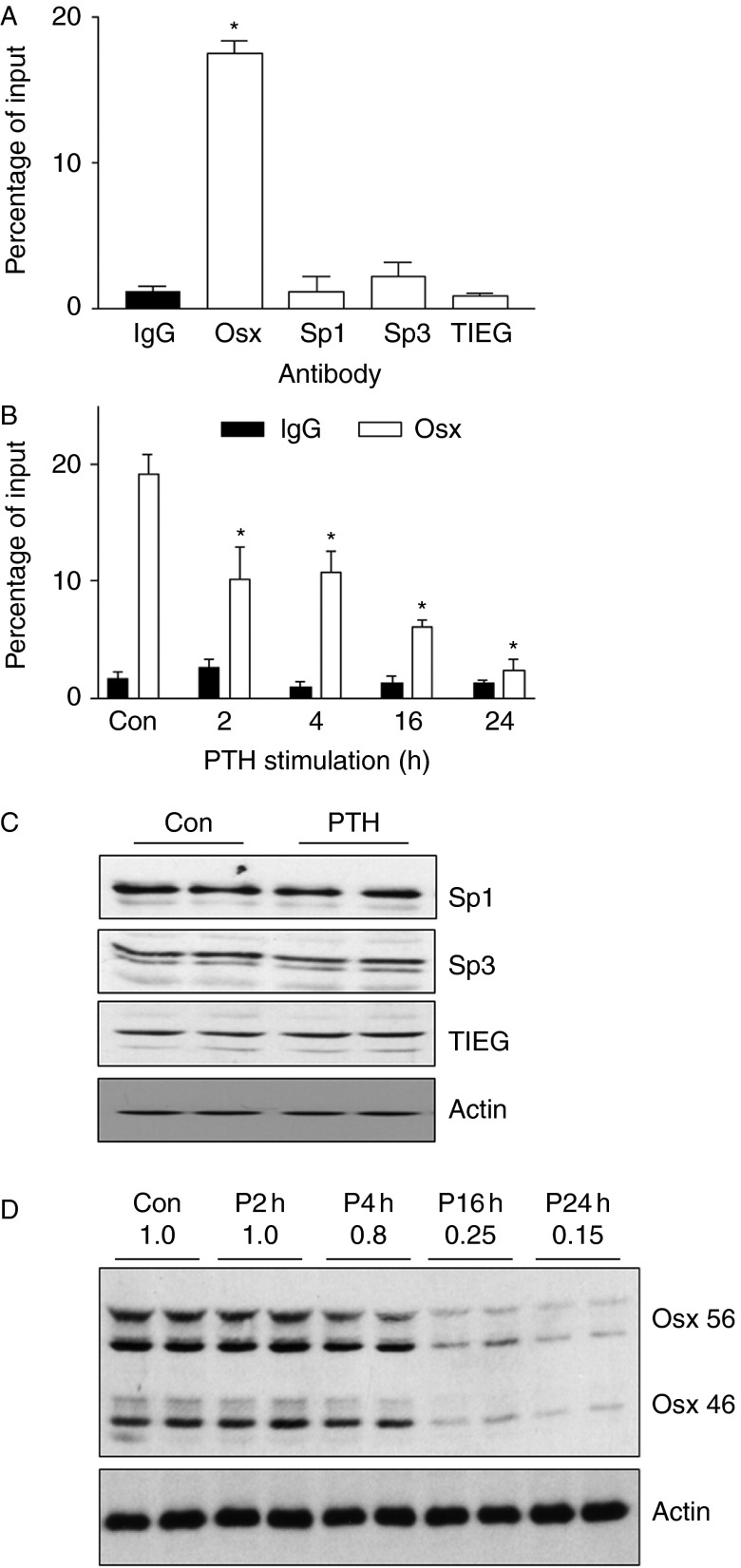
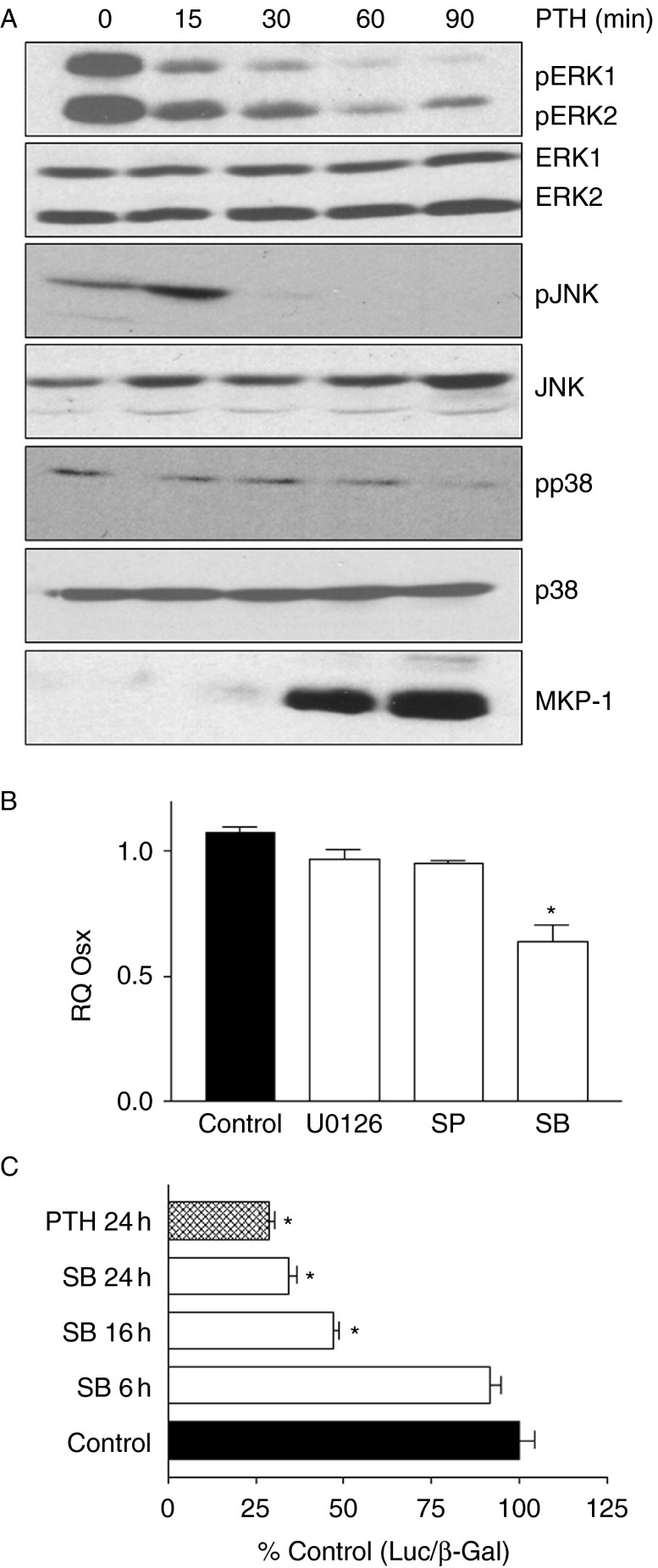
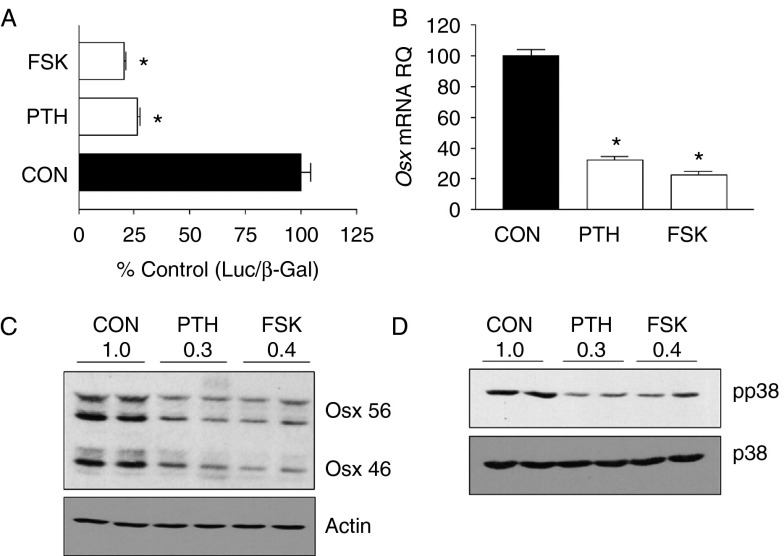
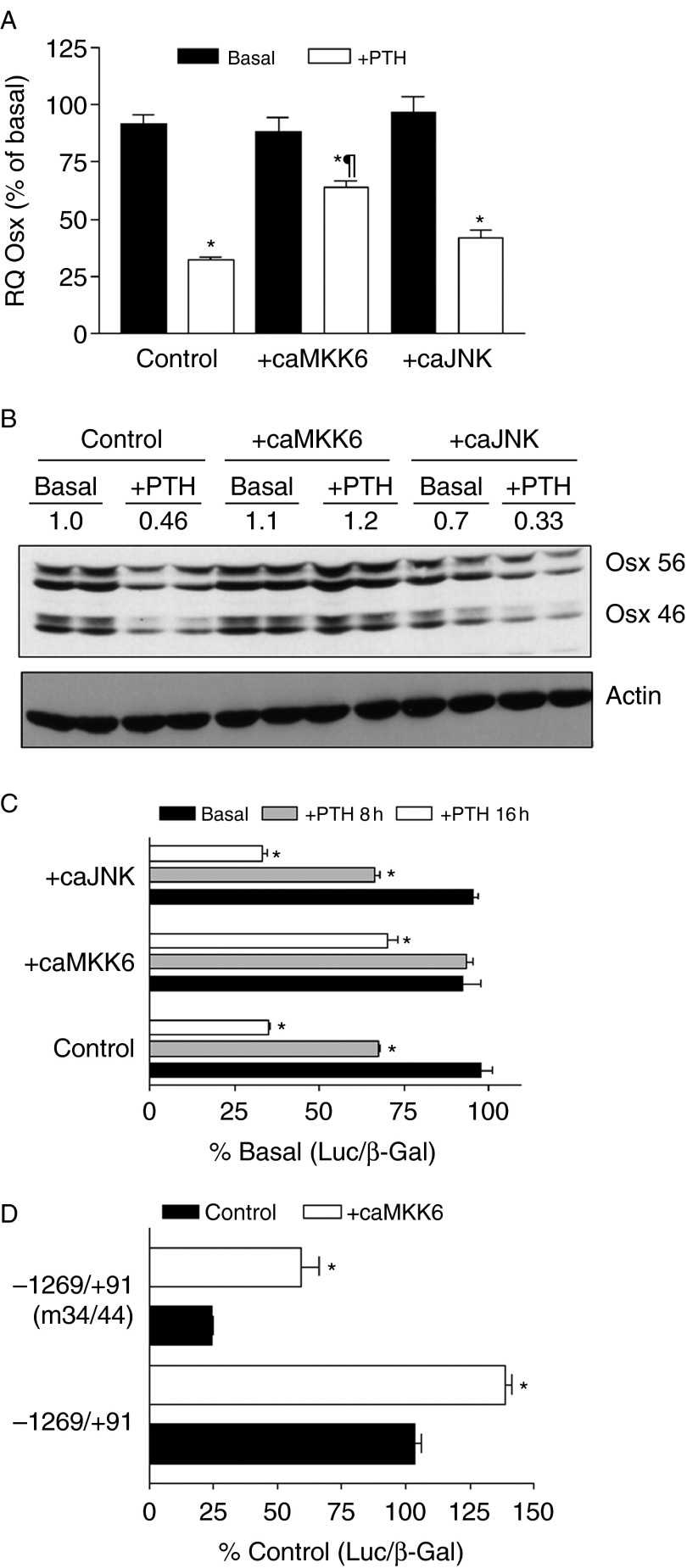
Osterix (Osx, Sp7) is a zinc-finger transcription factor belonging to the specificity protein (Sp) family expressed in cells of the osteoblast lineage in the developing skeleton where it regulates expression of a number of osteoblastic genes. We previously reported inhibition of osterix mRNA and protein by parathyroid hormone (PTH) stimulation of cAMP in osteoblasts. We here show that Osx expression in osteoblasts is regulated by Sp proteins as demonstrated by mithramycin A inhibition of Osx mRNA and OSX protein levels. Mutation of putative transcription factor binding sites within the Osx promoter demonstrated a tandem repeat sequence that selectively binds OSX but not other Sp factors expressed in osteoblasts (Sp1, Sp3, or Tieg (Klf10)). Mutation of either or both the repeat sequences inhibited 90% of the promoter activity and also abrogated some of the PTH-mediated inhibition of the promoter. Previous studies have shown growth factor regulation of Osx expression by MAPK proteins, particularly p38 phosphorylation of OSX that increases its transcriptional activity. PTH stimulation of osteoblasts inhibits MAPK components (ERK, JNK, and p38) but inhibition of Osx mRNA and protein expression by PTH was selectively mimicked by p38 inhibition and expression of constitutively active MKK6, which stimulates p38, blocked PTH inhibition of OSX. Together, our studies suggest that OSX autoregulation is a major mechanism in osteoblasts and that PTH stimulation inhibits osterix by inhibition of p38 MAPK regulation of OSX.
Keywords: Sp factor; autoregulation; osteoblasts; osterix; parathyroid hormone.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
