

Mechanism of Siglec-8-mediated cell death in IL-5-activated eosinophils: role for reactive oxygen species-enhanced MEK/ERK activation
- PMID: 23684072
- PMCID: PMC4042061
- DOI: 10.1016/j.jaci.2013.03.024
Mechanism of Siglec-8-mediated cell death in IL-5-activated eosinophils: role for reactive oxygen species-enhanced MEK/ERK activation
Abstract
Background: Sialic acid-binding immunoglobulin-like lectin (Siglec)-8 is expressed on human eosinophils, where its ligation induces cell death. Paradoxically, Siglec-8-mediated cell death is markedly enhanced by the presence of the activation and survival factor IL-5 and becomes independent of caspase activity.
Objective: In this report we investigate the mechanism of Siglec-8-mediated cell death in activated eosinophils.
Methods: Human peripheral blood eosinophils were treated with agonistic anti-Siglec-8 antibody and IL-5, and cell death was determined by using flow cytometry and morphology. Phosphorylation of mitogen-activated protein kinase (MAPK) was determined by using phosphoLuminex, flow cytometry, and Western blotting. Reactive oxygen species (ROS) accumulation was determined by using dihydrorhodamine fluorescence.
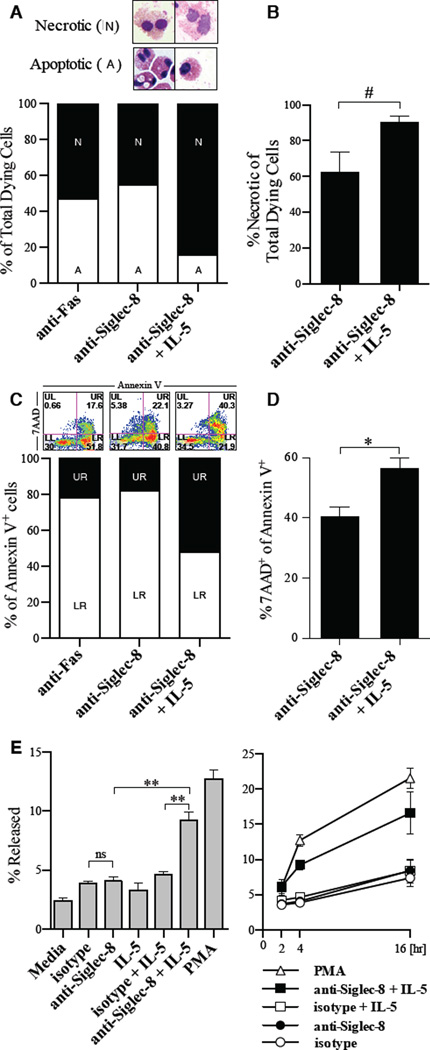
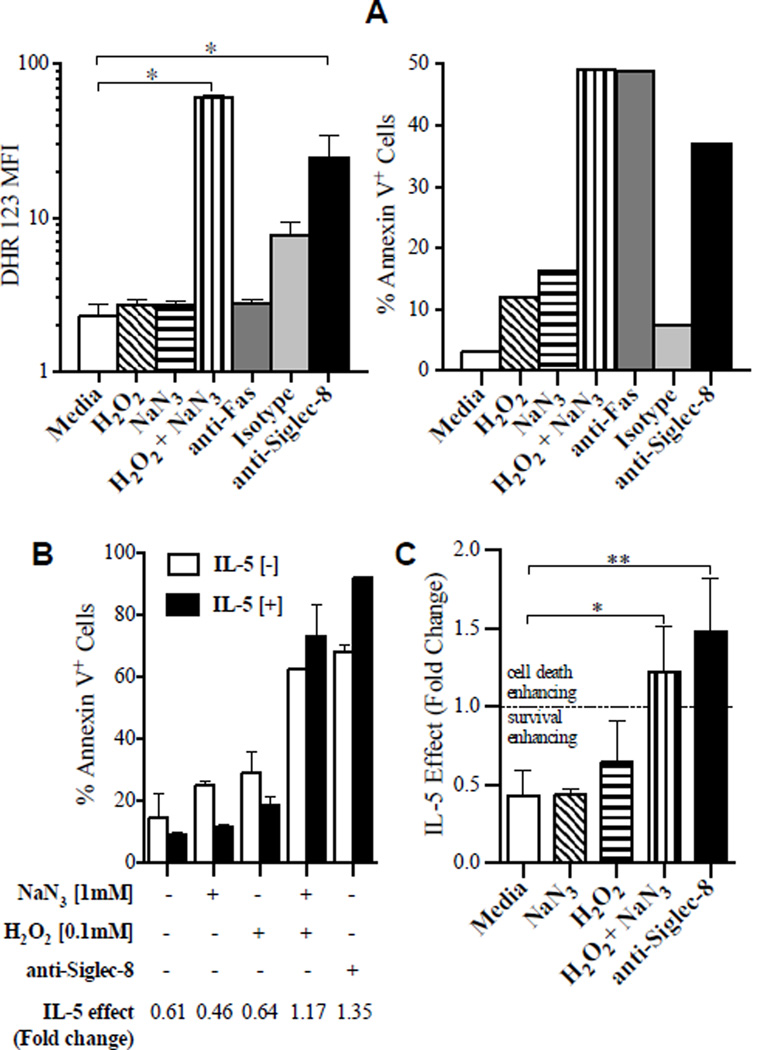
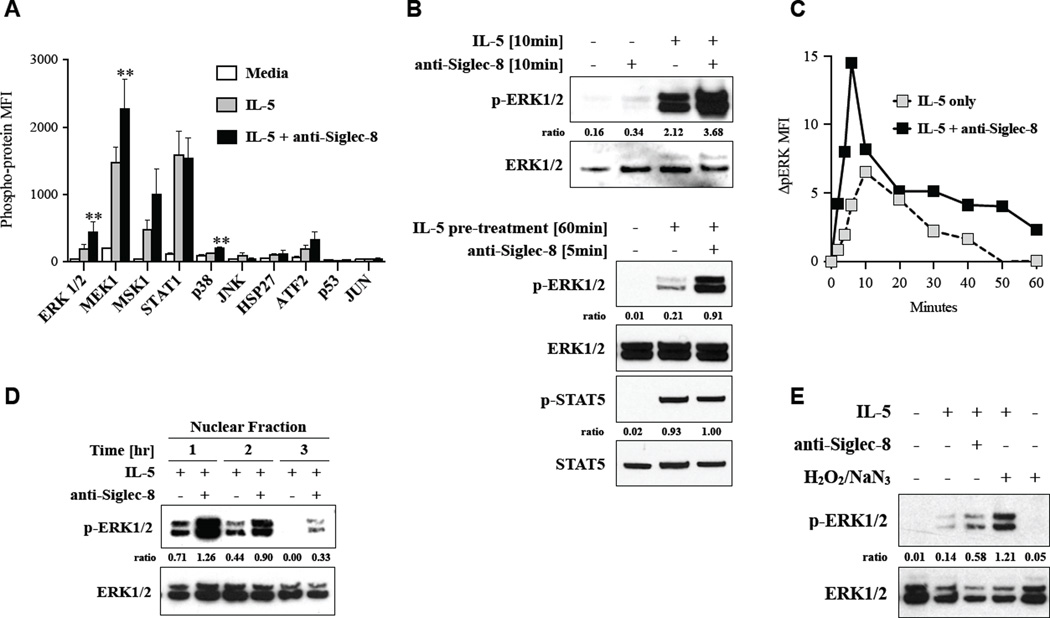
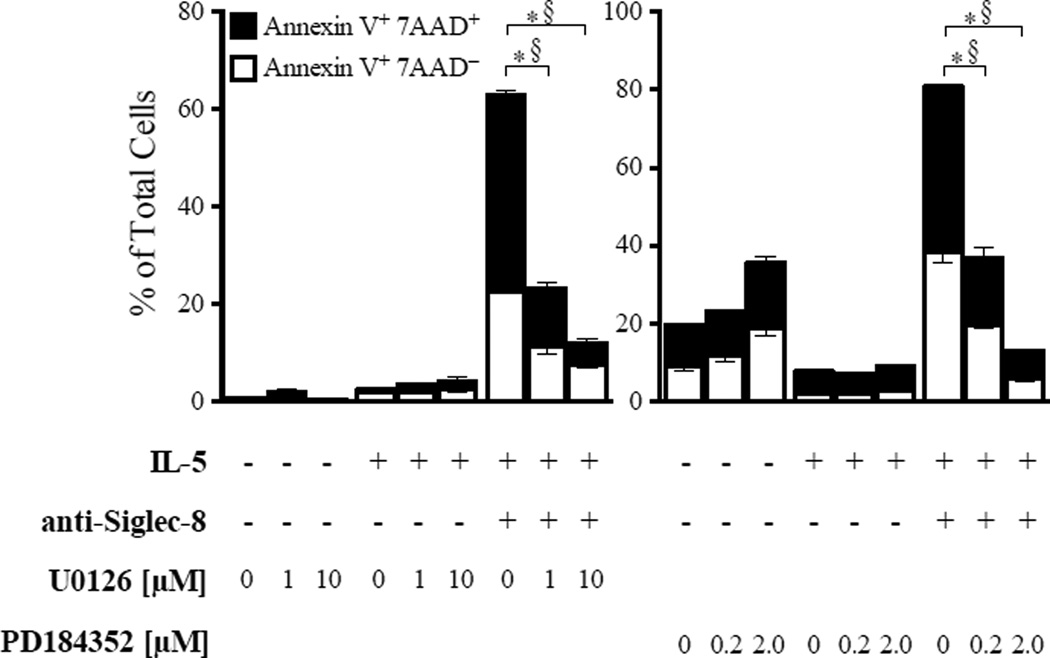
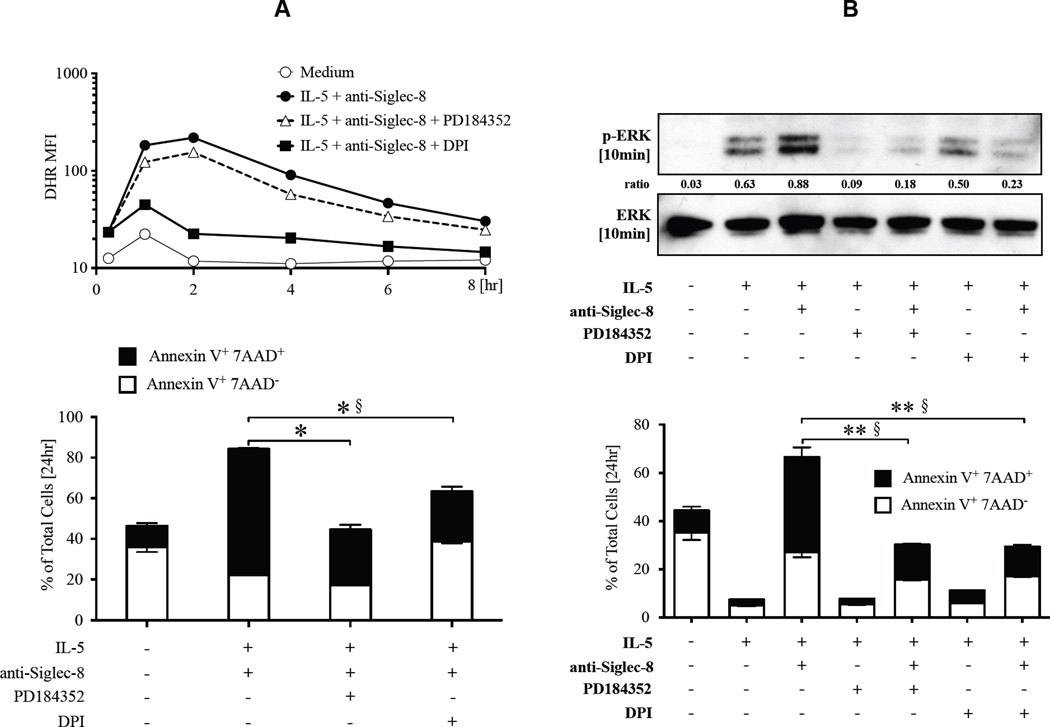
Results: Costimulation with anti-Siglec-8 and IL-5 significantly increased the rate and proportion of cell death by means of necrosis accompanied by granule release compared with that seen after stimulation with anti-Siglec-8 alone, in which apoptosis predominated. Together with the caspase-independent mode of cell death in costimulated cells, these findings suggest the activation of a specific and distinct biochemical pathway of cell death during anti-Siglec-8/IL-5 costimulation. Phosphorylation of extracellular signal-regulated kinase (ERK) 1/2 and MAPK-ERK kinase (MEK) 1 was significantly enhanced and sustained in costimulated cells compared with that seen in cells stimulated with IL-5 alone; anti-Siglec-8 alone did not cause ERK1/2 phosphorylation. MEK1 inhibitors blocked anti-Siglec-8/IL-5-induced cell death. ROS accumulation was induced by Siglec-8 ligation in a MEK-independent manner. In contrast, an ROS inhibitor prevented the anti-Siglec-8/IL-5-induced enhancement of ERK phosphorylation and cell death. Exogenous ROS mimicked stimulation by anti-Siglec-8 and was sufficient to induce enhanced cell death in IL-5-treated cells. Collectively, these data suggest that the enhancement of ERK phosphorylation is downstream of ROS generation.
Conclusions: In activated eosinophils ligation of Siglec-8 leads to ROS-dependent enhancement of IL-5-induced ERK phosphorylation, which results in a novel mode of biochemically regulated eosinophil cell death.
Keywords: 7-Aminoactinomycin D; 7AAD; DPI; DUSP; Diphenyleneiodonium; Dual-specificity phosphatase; EPX; ERK; Eosinophil peroxidase; Eosinophils; Extracellular signal-regulated kinase; H(2)O(2); HRP; Horseradish peroxidase; Hydrogen peroxide; JNK; MAPK; MAPK-ERK kinase; MEK; Mitogen-activated protein kinase; NaN(3); PFA; Paraformaldehyde; ROS; Reactive oxygen species; STAT; Sialic acid–binding immunoglobulin-like lectin; Siglec; Signal transducer and activator of transcription; Sodium azide; c-Jun N-terminal kinase; cell death; signaling.
Copyright © 2013 American Academy of Allergy, Asthma & Immunology. Published by Mosby, Inc. All rights reserved.
Conflict of interest statement
Figures
Comment in
-
Lysis of primed eosinophils in severe asthma.J Allergy Clin Immunol. 2013 Dec;132(6):1459-60. doi: 10.1016/j.jaci.2013.09.036. Epub 2013 Nov 1. J Allergy Clin Immunol. 2013. PMID: 24184146 No abstract available.
References
-
- Floyd H, Ni J, Cornish AL, Zeng Z, Liu D, Carter KC, et al. Siglec-8. A novel eosinophil-specific member of the immunoglobulin superfamily. J Biol Chem. 2000;275:861–866. - PubMed
-
- Nutku E, Aizawa H, Hudson SA, Bochner BS. Ligation of Siglec-8: a selective mechanism for induction of human eosinophil apoptosis. Blood. 2003;101:5014–5020. - PubMed
-
- Kikly KK, Bochner BS, Freeman SD, Tan KB, Gallagher KT, D'Alessio KJ, et al. Identification of SAF-2, a novel siglec expressed on eosinophils, mast cells, and basophils. J Allergy Clin Immunol. 2000;105:1093–1100. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
