

Molecular dissection of step 2 catalysis of yeast pre-mRNA splicing investigated in a purified system
- PMID: 23685439
- PMCID: PMC3683925
- DOI: 10.1261/rna.039024.113
Molecular dissection of step 2 catalysis of yeast pre-mRNA splicing investigated in a purified system
Abstract
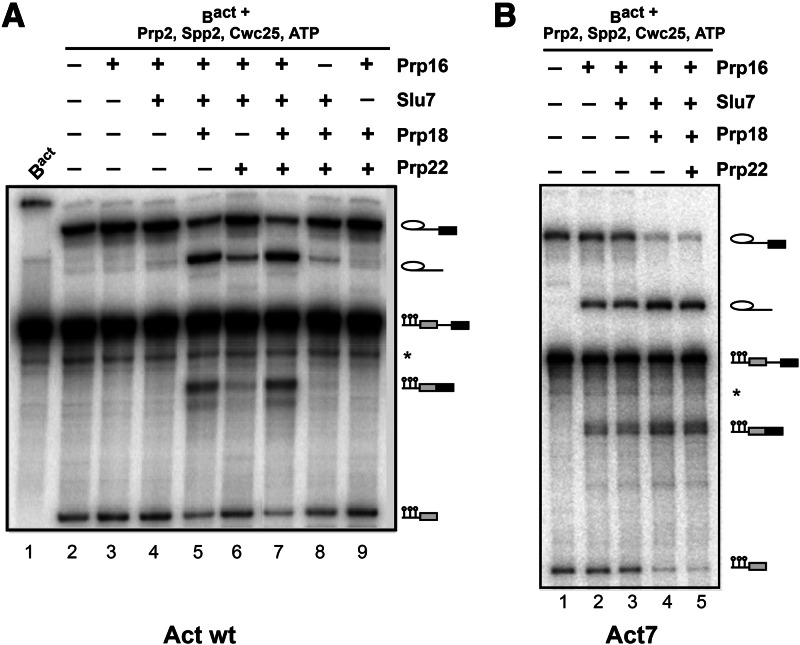
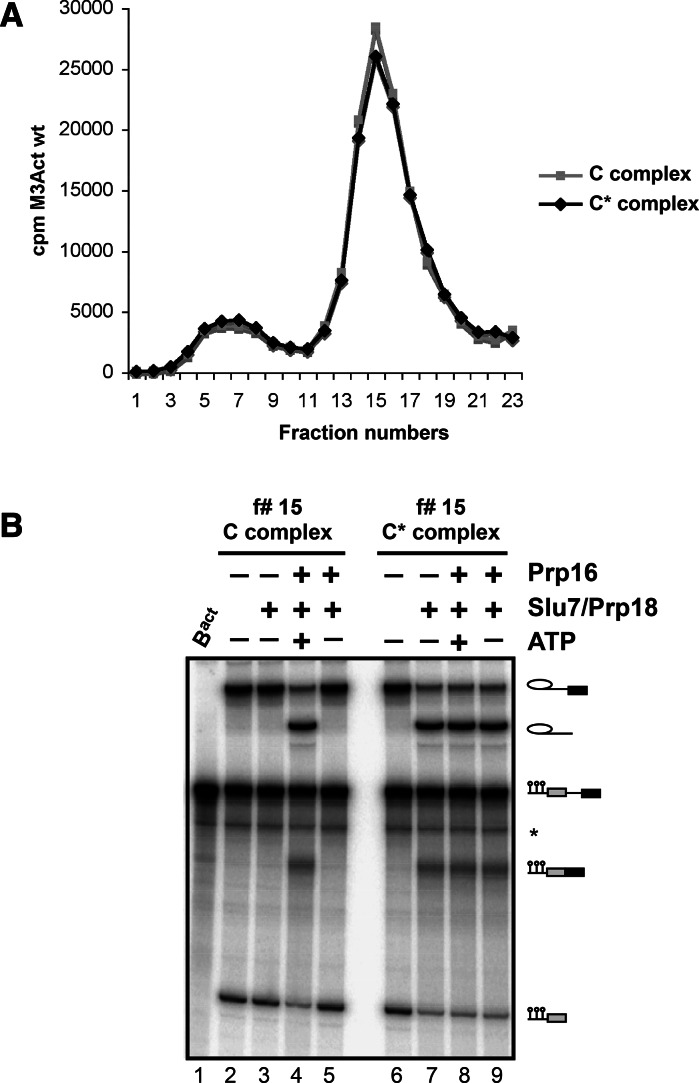
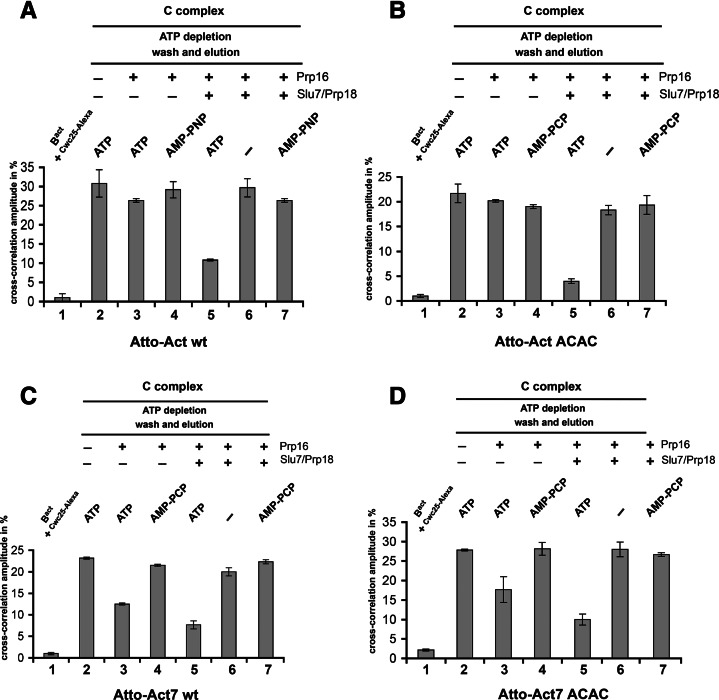
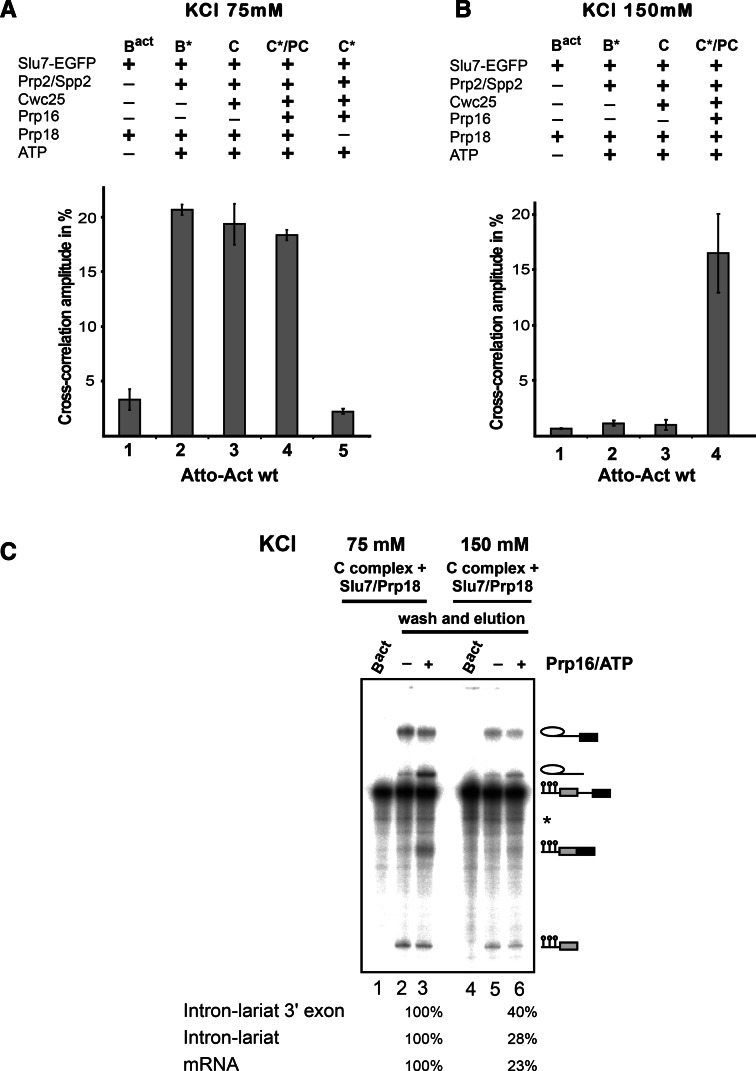
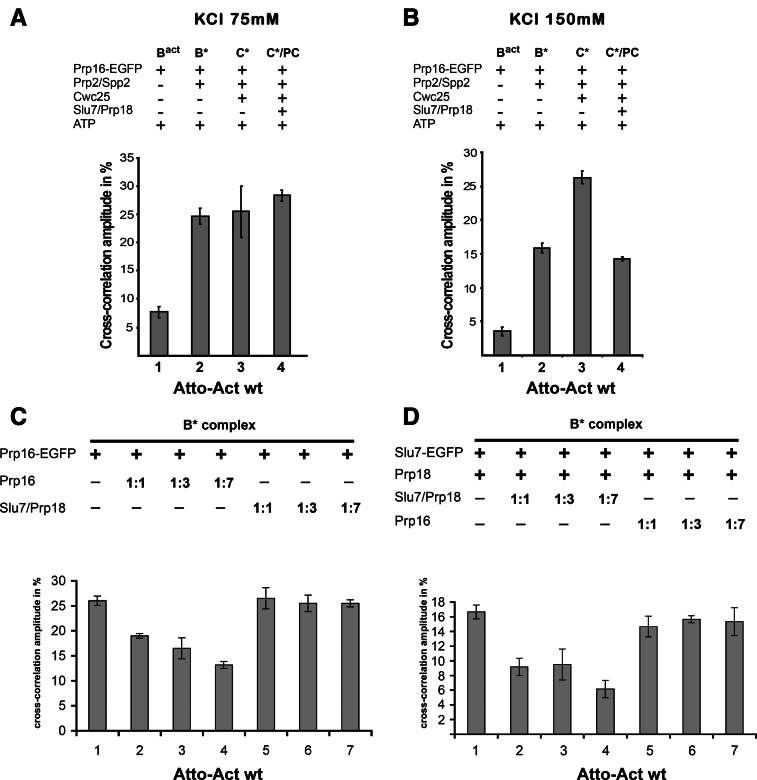
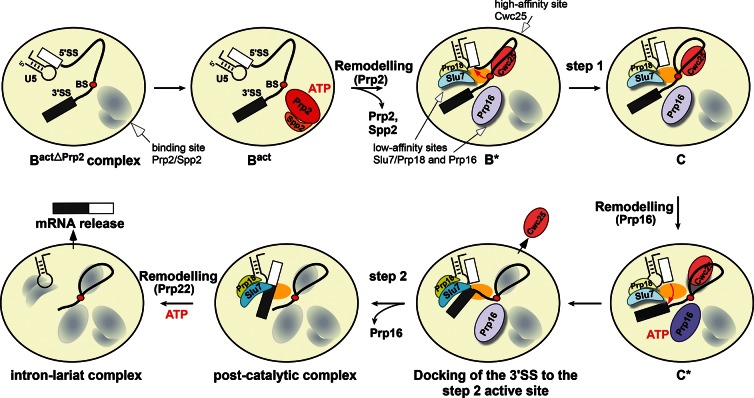
Step 2 catalysis of pre-mRNA splicing entails the excision of the intron and ligation of the 5' and 3' exons. The tasks of the splicing factors Prp16, Slu7, Prp18, and Prp22 in the formation of the step 2 active site of the spliceosome and in exon ligation, and the timing of their recruitment, remain poorly understood. Using a purified yeast in vitro splicing system, we show that only the DEAH-box ATPase Prp16 is required for formation of a functional step 2 active site and for exon ligation. Efficient docking of the 3' splice site (3'SS) to the active site requires only Slu7/Prp18 but not Prp22. Spliceosome remodeling by Prp16 appears to be subtle as only the step 1 factor Cwc25 is dissociated prior to step 2 catalysis, with its release dependent on docking of the 3'SS to the active site and Prp16 action. We show by fluorescence cross-correlation spectroscopy that Slu7/Prp18 and Prp16 bind early to distinct, low-affinity binding sites on the step-1-activated B* spliceosome, which are subsequently converted into high-affinity sites. Our results shed new light on the factor requirements for step 2 catalysis and the dynamics of step 1 and 2 factors during the catalytic steps of splicing.
Keywords: Prp22; Slu7 Prp18; spliceosome; step 2 catalysis; step 2 factors/Prp16.
Figures
References
-
- Burgess SM, Guthrie C 1993. A mechanism to enhance mRNA splicing fidelity: The RNA-dependent ATPase Prp16 governs usage of a discard pathway for aberrant lariat intermediates. Cell 73: 1377–1391 - PubMed
-
- Burgess S, Couto JR, Guthrie C 1990. A putative ATP binding protein influences the fidelity of branchpoint recognition in yeast splicing. Cell 60: 705–717 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases