

Deletions of recessive disease genes: CNV contribution to carrier states and disease-causing alleles
- PMID: 23685542
- PMCID: PMC3759716
- DOI: 10.1101/gr.156075.113
Deletions of recessive disease genes: CNV contribution to carrier states and disease-causing alleles
Abstract
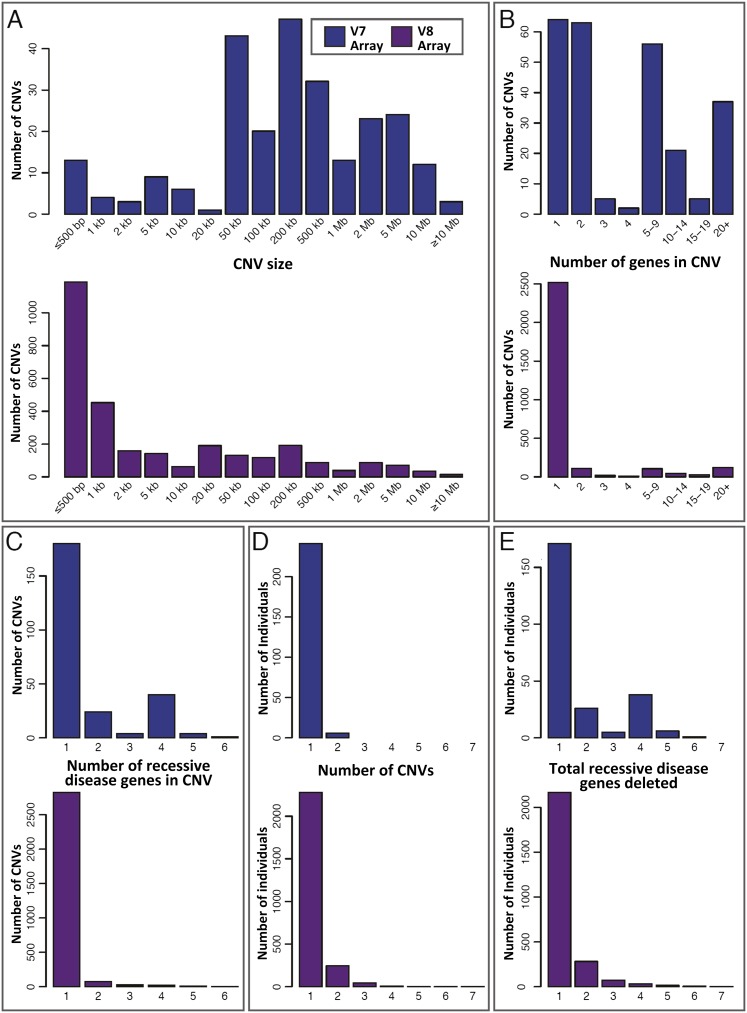
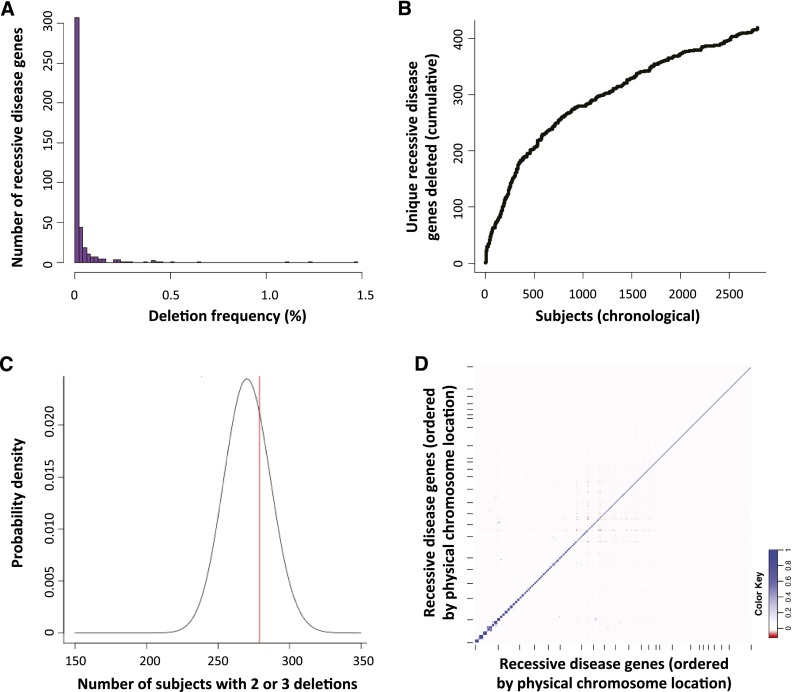
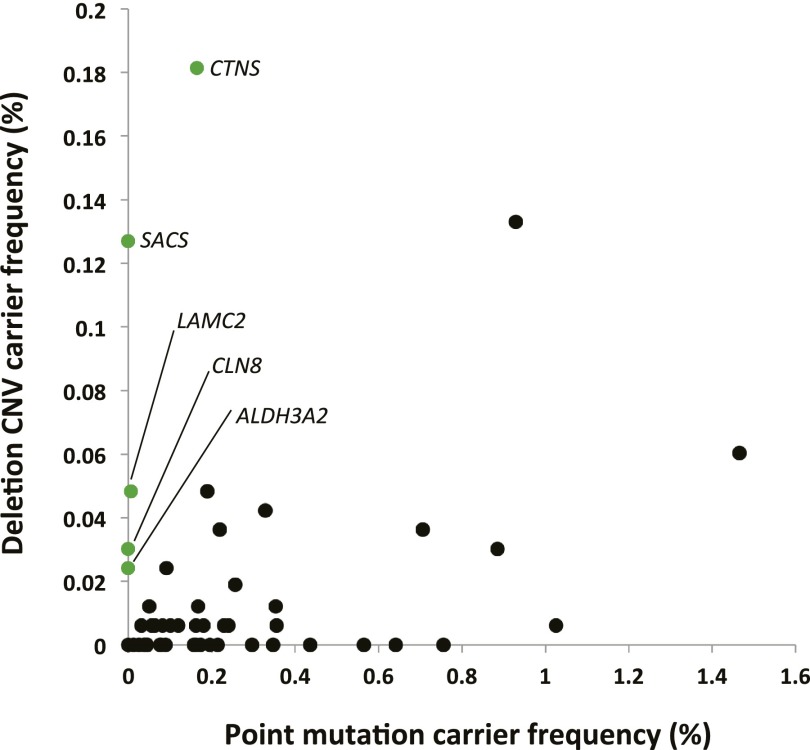
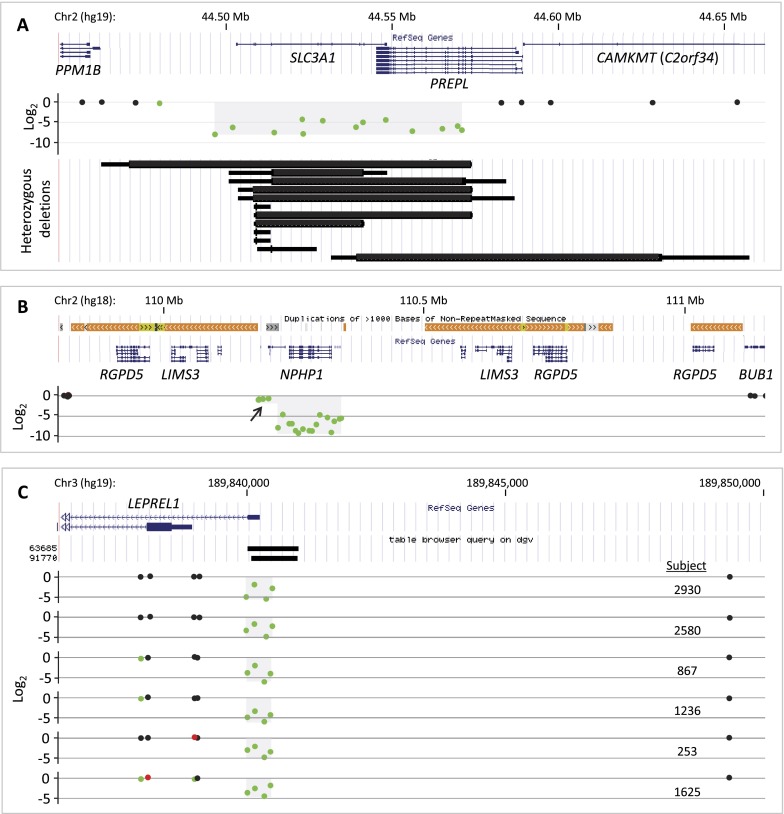
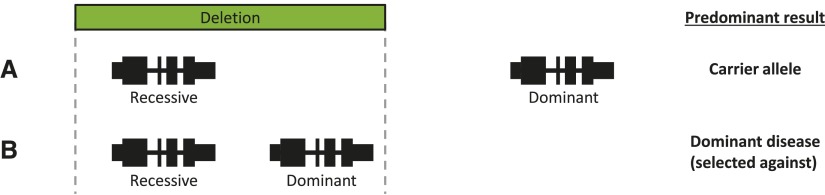
Over 1200 recessive disease genes have been described in humans. The prevalence, allelic architecture, and per-genome load of pathogenic alleles in these genes remain to be fully elucidated, as does the contribution of DNA copy-number variants (CNVs) to carrier status and recessive disease. We mined CNV data from 21,470 individuals obtained by array-comparative genomic hybridization in a clinical diagnostic setting to identify deletions encompassing or disrupting recessive disease genes. We identified 3212 heterozygous potential carrier deletions affecting 419 unique recessive disease genes. Deletion frequency of these genes ranged from one occurrence to 1.5%. When compared with recessive disease genes never deleted in our cohort, the 419 recessive disease genes affected by at least one carrier deletion were longer and located farther from known dominant disease genes, suggesting that the formation and/or prevalence of carrier CNVs may be affected by both local and adjacent genomic features and by selection. Some subjects had multiple carrier CNVs (307 subjects) and/or carrier deletions encompassing more than one recessive disease gene (206 deletions). Heterozygous deletions spanning multiple recessive disease genes may confer carrier status for multiple single-gene disorders, for complex syndromes resulting from the combination of two or more recessive conditions, or may potentially cause clinical phenotypes due to a multiply heterozygous state. In addition to carrier mutations, we identified homozygous and hemizygous deletions potentially causative for recessive disease. We provide further evidence that CNVs contribute to the allelic architecture of both carrier and recessive disease-causing mutations. Thus, a complete recessive carrier screening method or diagnostic test should detect CNV alleles.
Figures


References
-
- Bacino CA, Cheung S-W 2010. Introductory comments on special section–genomic microduplications: When adding may equal subtracting. Am J Med Genet A 152A: 1063–1065 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases