

Gamma delta T cells are activated by polysaccharide K (PSK) and contribute to the anti-tumor effect of PSK
- PMID: 23685781
- PMCID: PMC3727629
- DOI: 10.1007/s00262-013-1436-4
Gamma delta T cells are activated by polysaccharide K (PSK) and contribute to the anti-tumor effect of PSK
Abstract
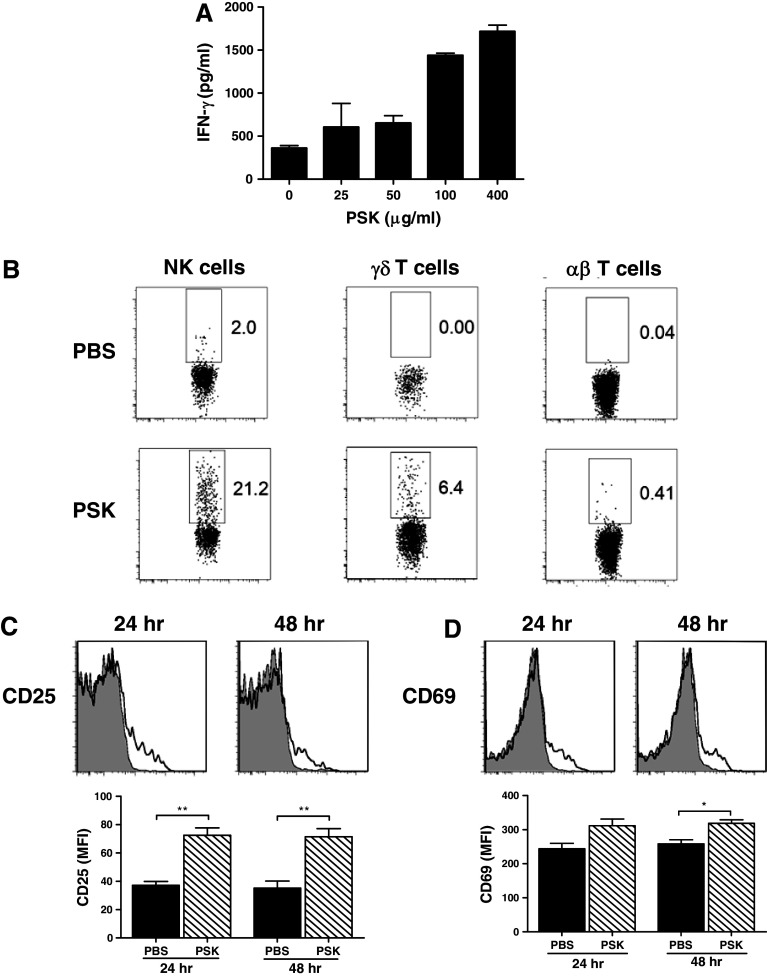
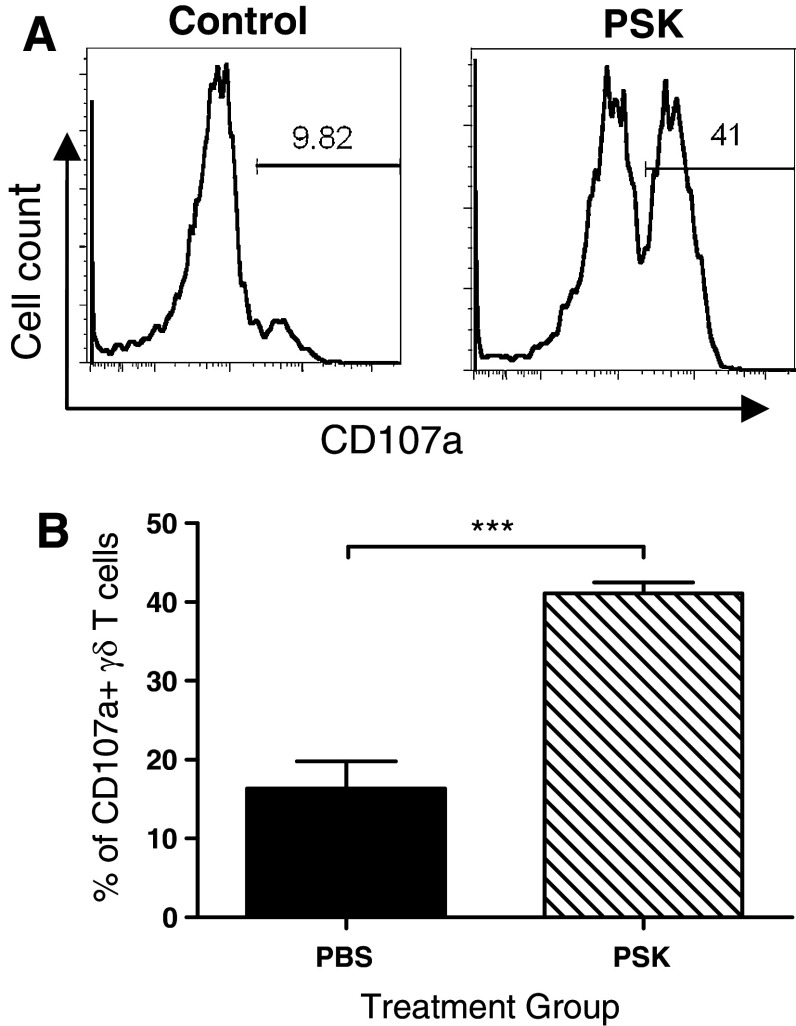
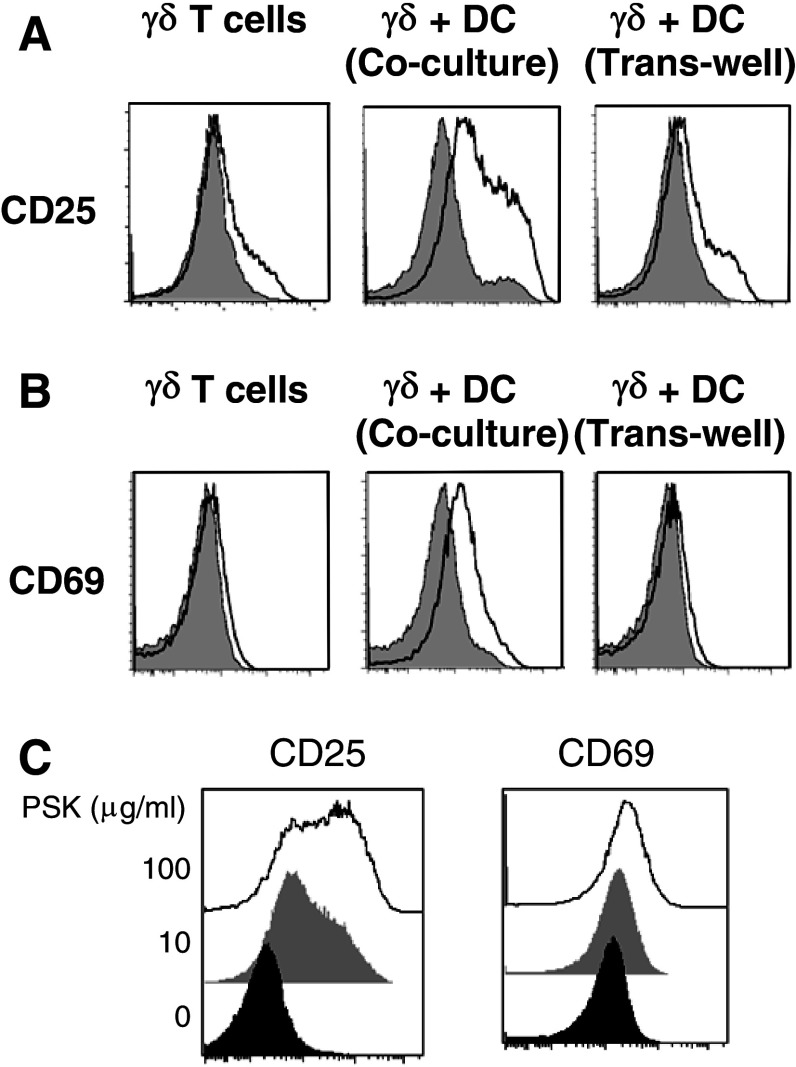
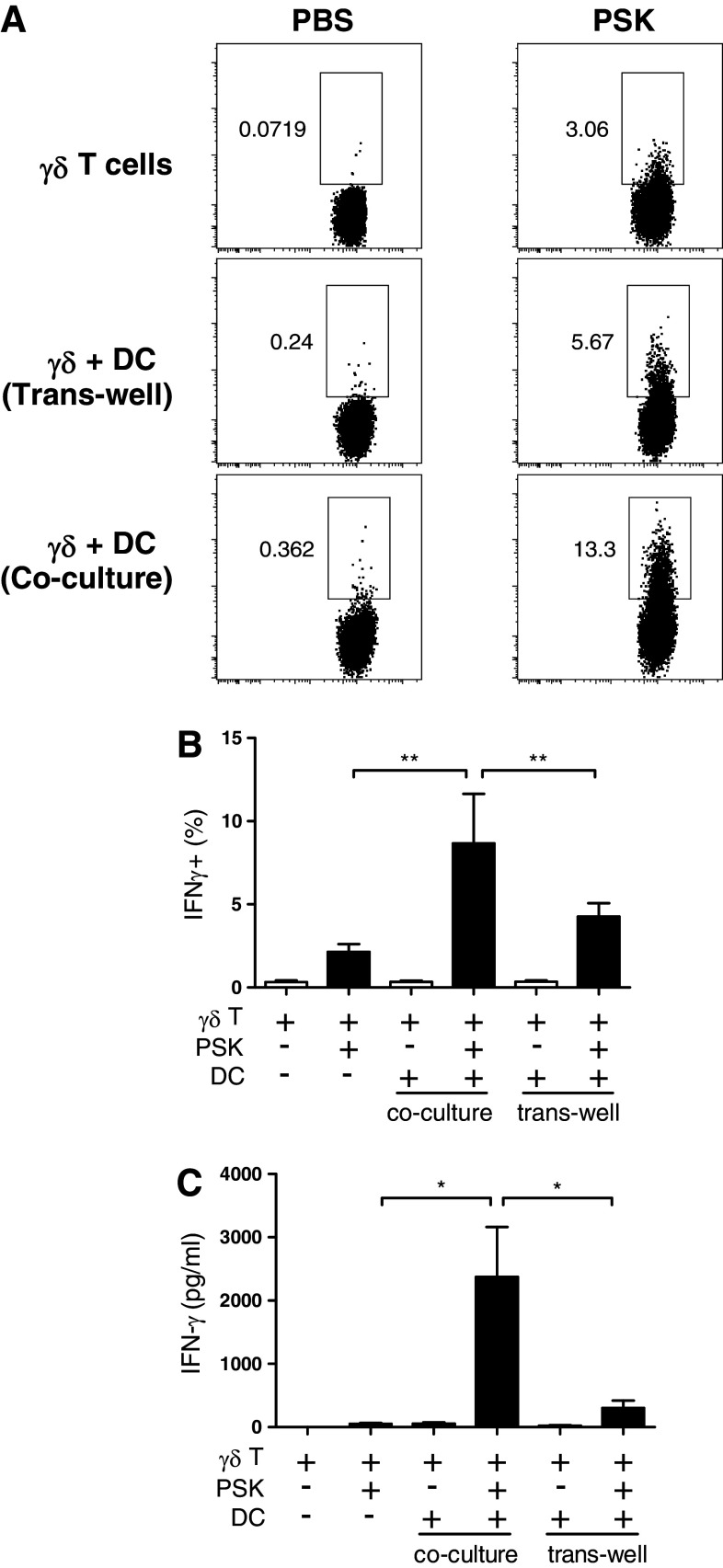
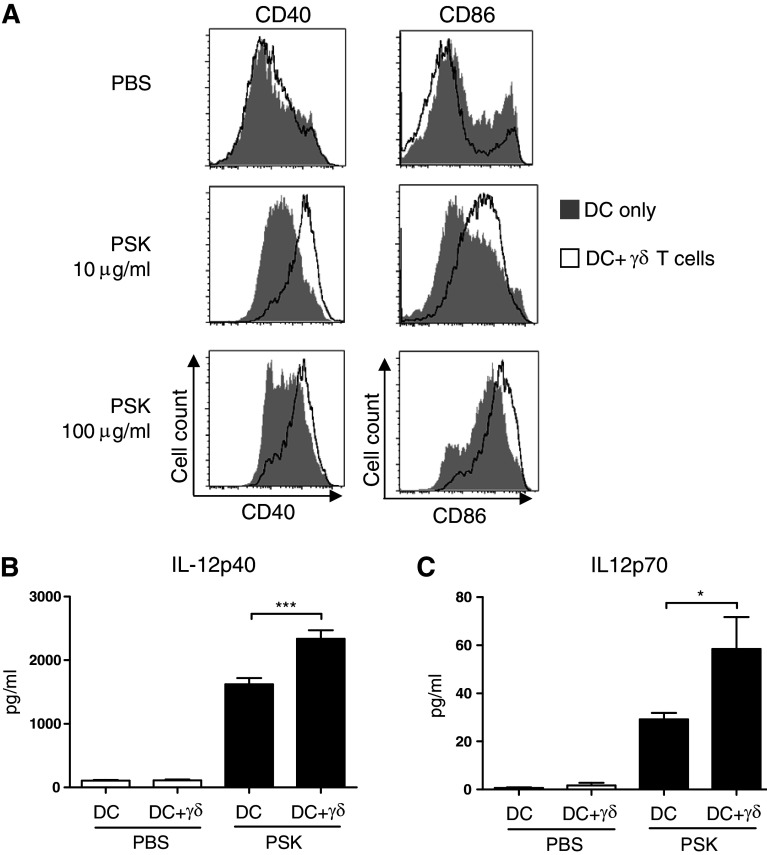
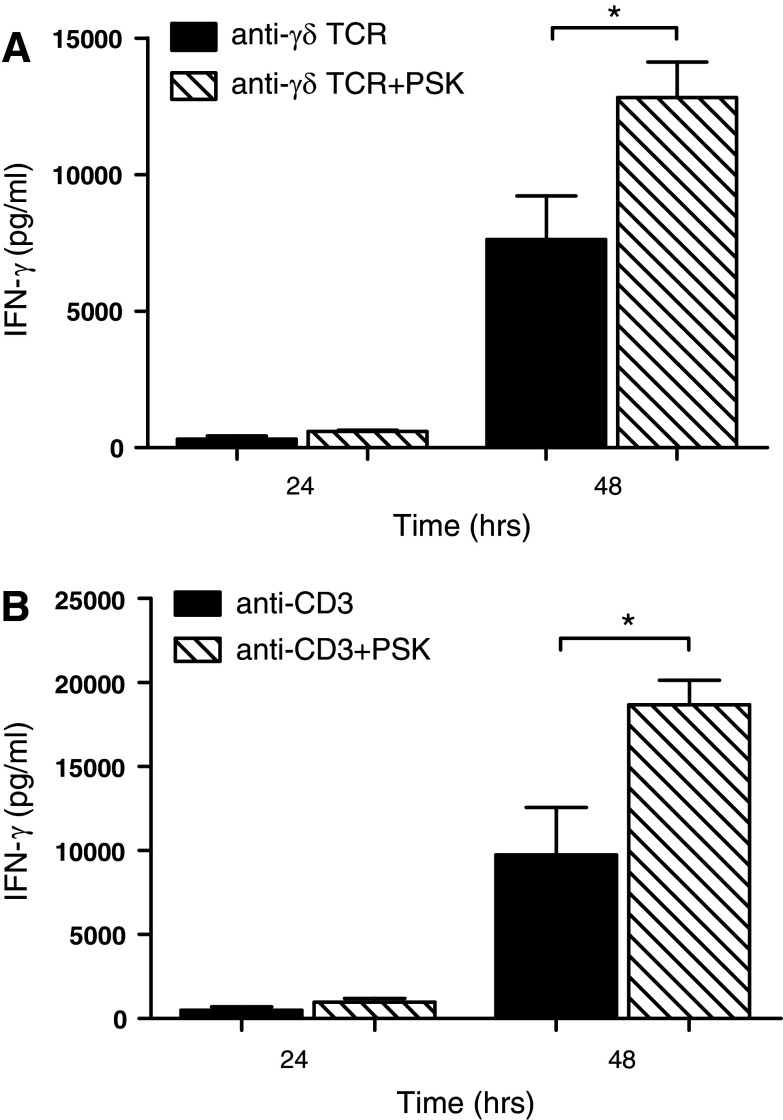
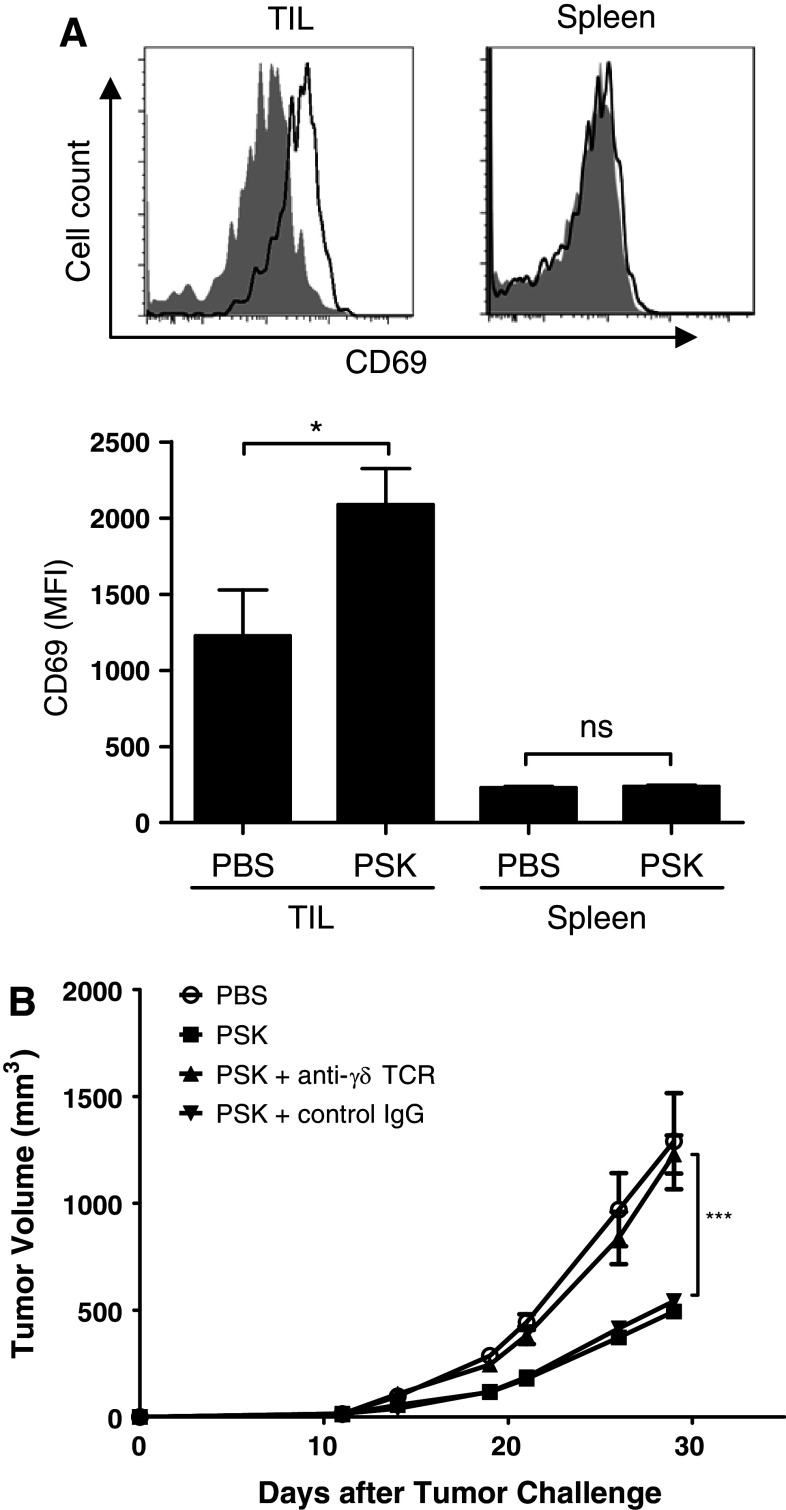
Polysaccharide K (PSK) is a widely used mushroom extract that has shown anti-tumor and immunomodulatory effects in both preclinical and clinical studies. Therefore, it is important to understand the mechanism of actions of PSK. We recently reported that PSK can activate toll-like receptor 2 and enhances the function of NK cells. The current study was undertaken to study the effect of PSK on gamma delta (γδ) T cells, another important arm of the innate immunity. In vitro experiments using mouse splenocytes showed that γδ T cells produce IFN-γ after treatment with PSK and have up-regulated expression of CD25, CD69, and CD107a. To investigate whether the effect of PSK on γδ T cells is direct or indirect, purified γδ T cells were cultured either alone or together with bone marrow-derived DC in a co-culture or trans-well system and then stimulated with PSK. Results showed that direct cell-to-cell contact between γδ T cells and DC is required for optimal activation of γδ T cells. There was also reciprocal activation of DC by PSK-activated γδ T cells, as demonstrated by higher expression of costimulatory molecules and enhanced production of IL-12 by DC in the presence of γδ T cells. PSK can also co-stimulate γδ T cells with anti-TCR and anti-CD3 stimulation, in the absence of DC. Finally, in vivo treatment with PSK activates γδ T cells among the tumor infiltrating lymphocytes, and depleting γδ T cells during PSK treatment attenuated the anti-tumor effect of PSK. All together, these results demonstrated that γδ T cells are activated by PSK and contribute to the anti-tumor effect of PSK.
Conflict of interest statement
The authors declare they have no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
