

N-truncated amyloid β (Aβ) 4-42 forms stable aggregates and induces acute and long-lasting behavioral deficits
- PMID: 23685882
- PMCID: PMC3722453
- DOI: 10.1007/s00401-013-1129-2
N-truncated amyloid β (Aβ) 4-42 forms stable aggregates and induces acute and long-lasting behavioral deficits
Abstract
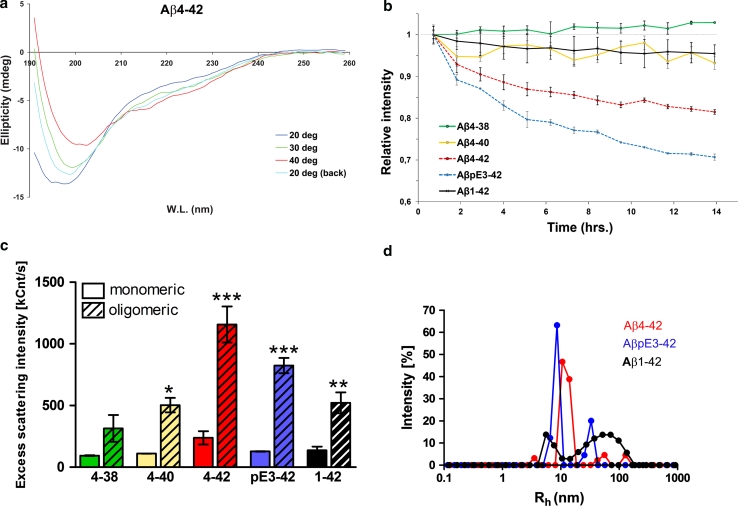
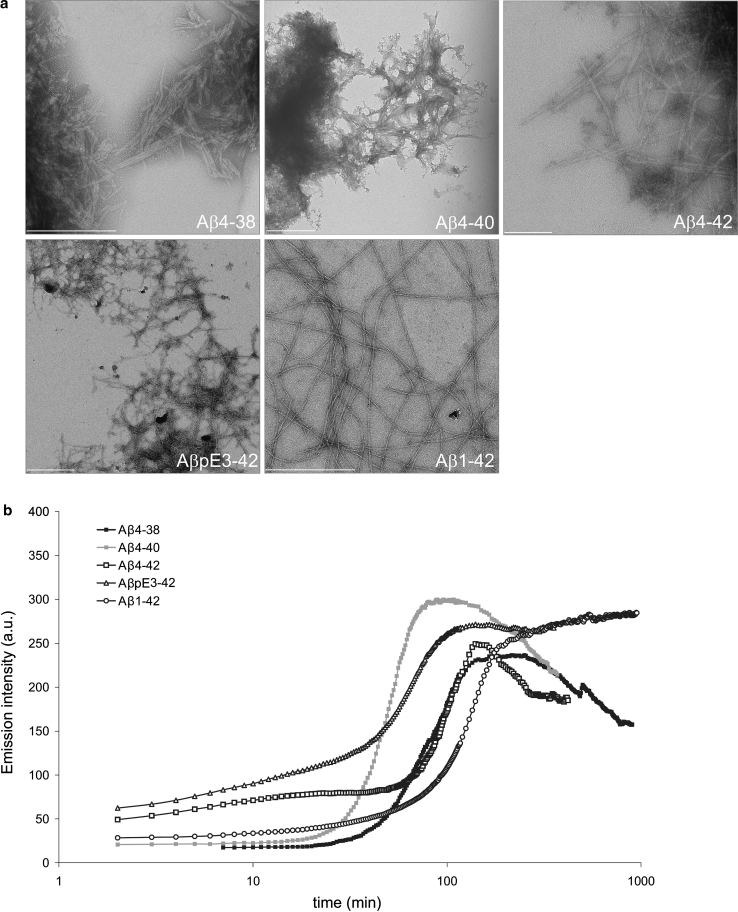
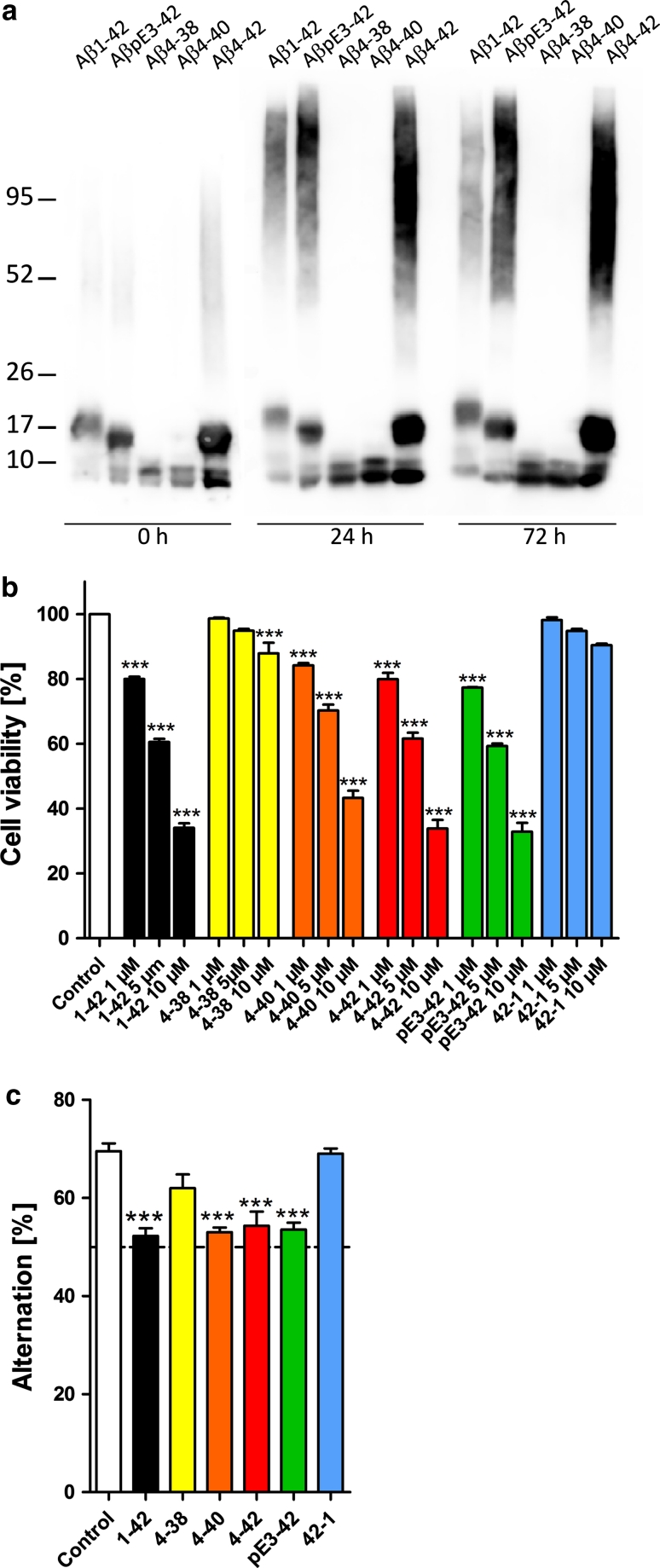
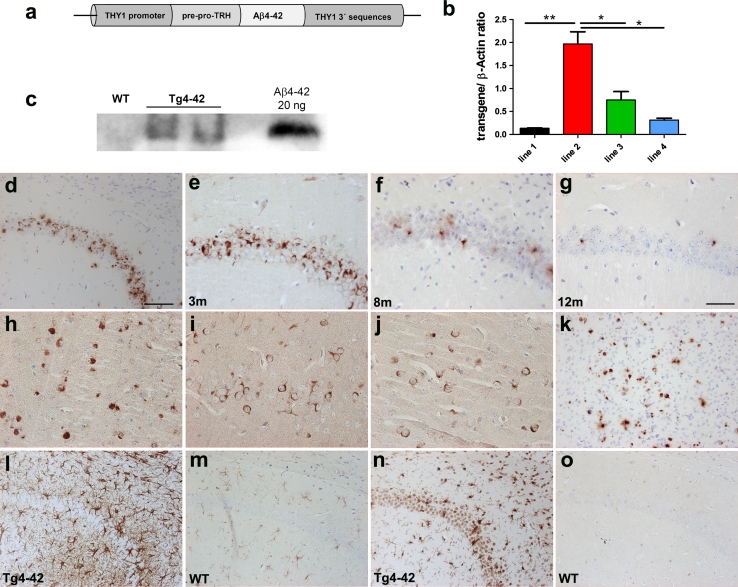
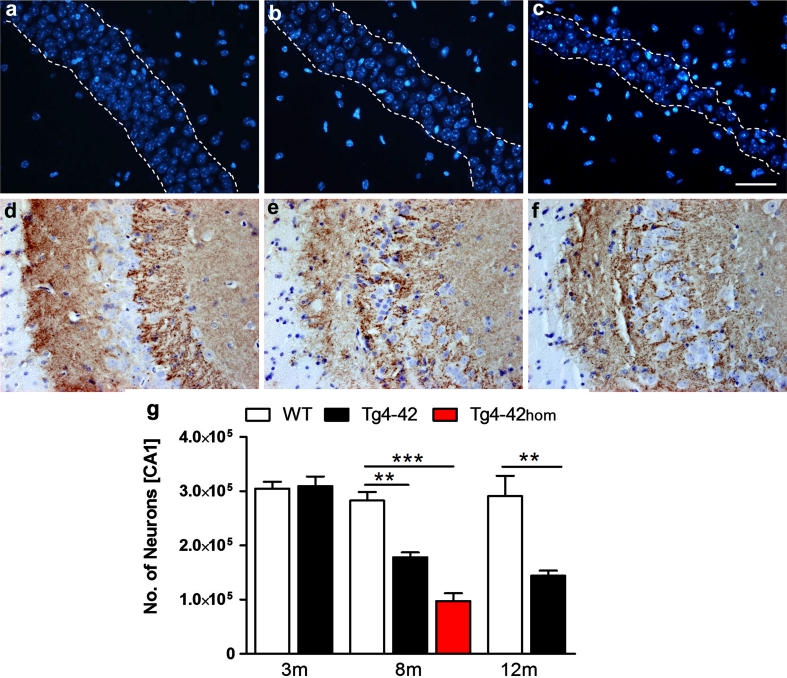
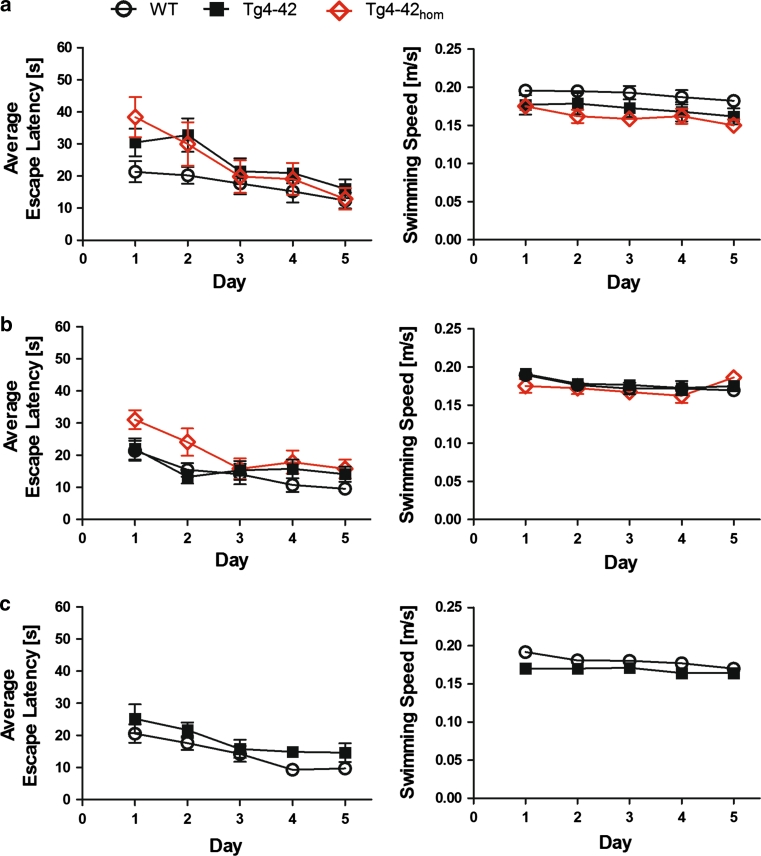
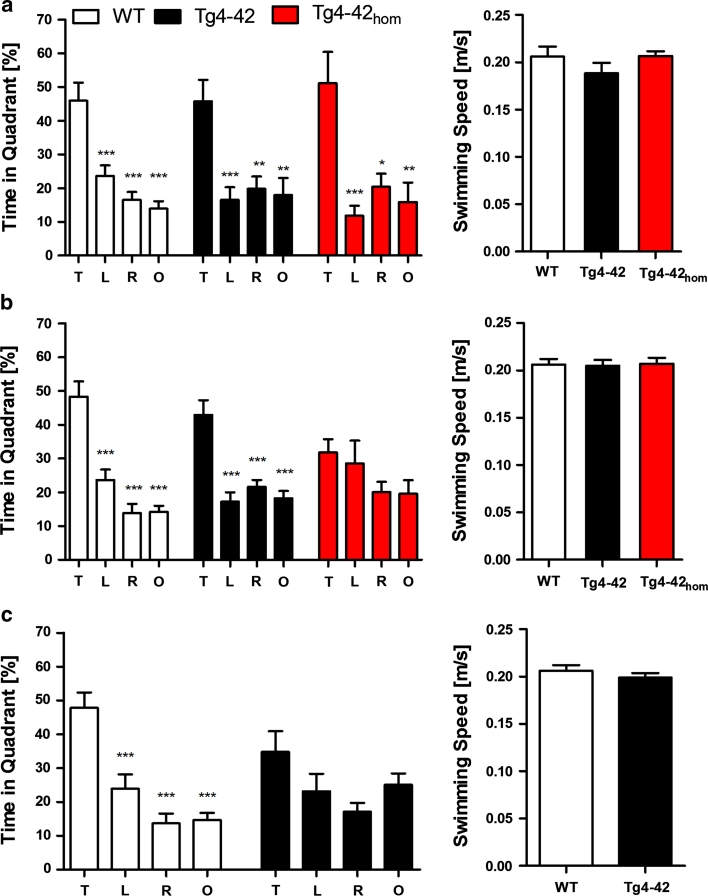
N-truncated Aβ4-42 is highly abundant in Alzheimer disease (AD) brain and was the first Aβ peptide discovered in AD plaques. However, a possible role in AD aetiology has largely been neglected. In the present report, we demonstrate that Aβ4-42 rapidly forms aggregates possessing a high aggregation propensity in terms of monomer consumption and oligomer formation. Short-term treatment of primary cortical neurons indicated that Aβ4-42 is as toxic as pyroglutamate Aβ3-42 and Aβ1-42. In line with these findings, treatment of wildtype mice using intraventricular Aβ injection induced significant working memory deficits with Aβ4-42, pyroglutamate Aβ3-42 and Aβ1-42. Transgenic mice expressing Aβ4-42 (Tg4-42 transgenic line) developed a massive CA1 pyramidal neuron loss in the hippocampus. The hippocampus-specific expression of Aβ4-42 correlates well with age-dependent spatial reference memory deficits assessed by the Morris water maze test. Our findings indicate that N-truncated Aβ4-42 triggers acute and long-lasting behavioral deficits comparable to AD typical memory dysfunction.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
