

Evidence for a common progenitor of epithelial and mesenchymal components of the liver
- PMID: 23686136
- PMCID: PMC3705603
- DOI: 10.1038/cdd.2013.49
Evidence for a common progenitor of epithelial and mesenchymal components of the liver
Abstract
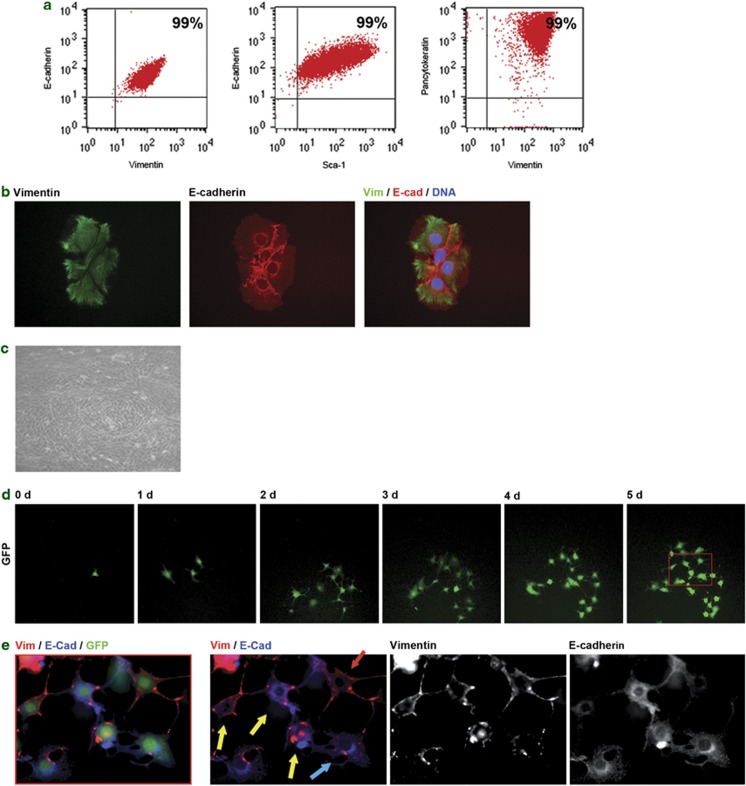
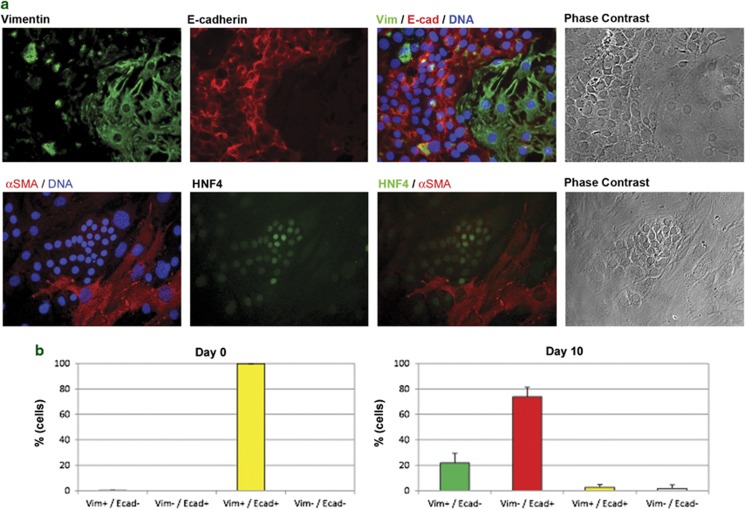
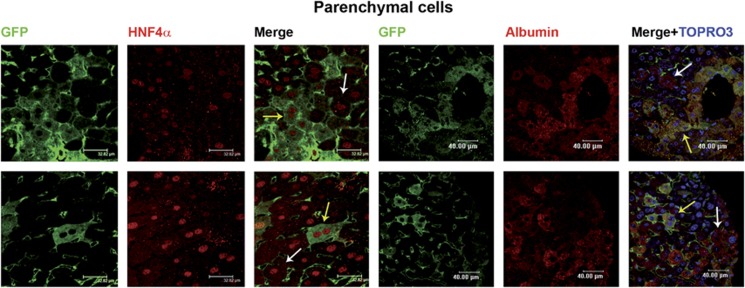
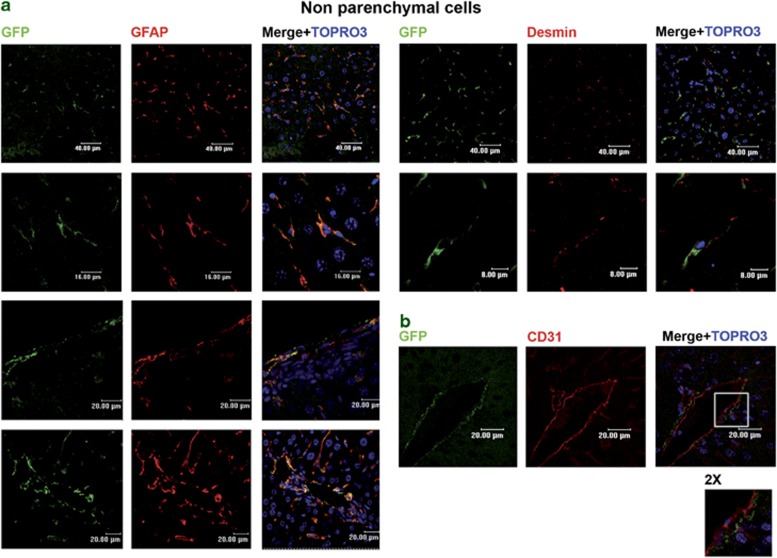
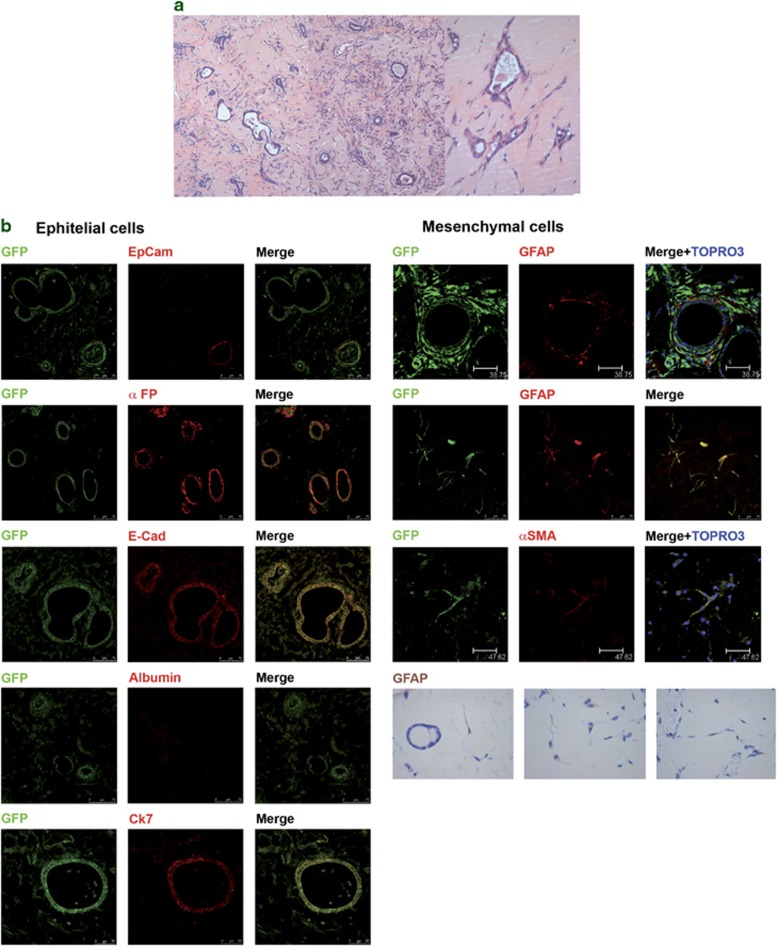
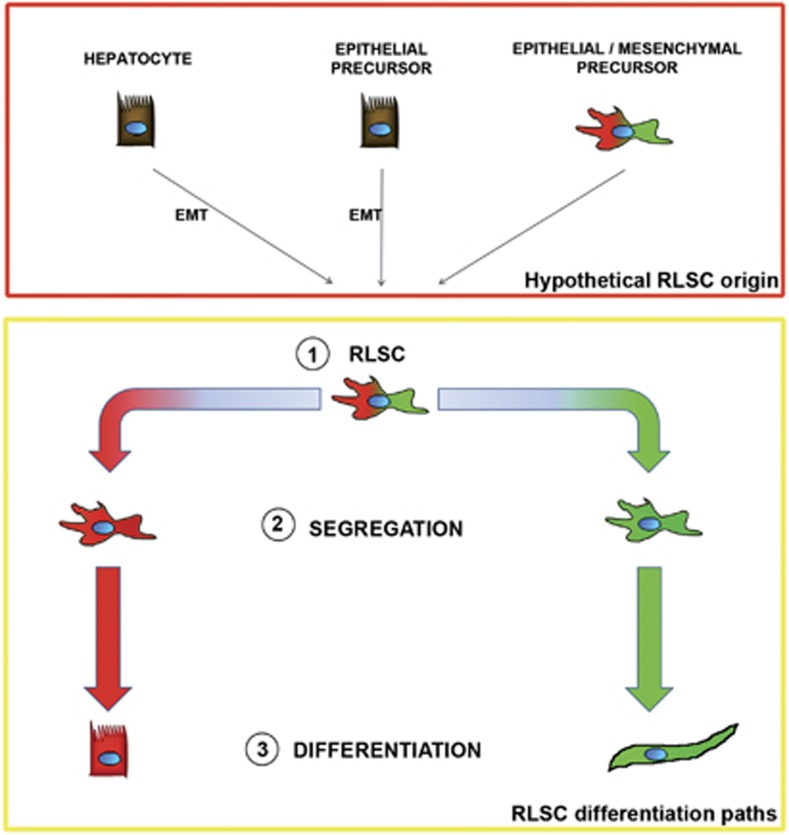
Tissues of the adult organism maintain the homeostasis and respond to injury by means of progenitor/stem cell compartments capable to give rise to appropriate progeny. In organs composed by histotypes of different embryological origins (e.g. the liver), the tissue turnover may in theory involve different stem/precursor cells able to respond coordinately to physiological or pathological stimuli. In the liver, a progenitor cell compartment, giving rise to hepatocytes and cholangiocytes, can be activated by chronic injury inhibiting hepatocyte proliferation. The precursor compartment guaranteeing turnover of hepatic stellate cells (HSCs) (perisinusoidal cells implicated with the origin of the liver fibrosis) in adult organ is yet unveiled. We show here that epithelial and mesenchymal liver cells (hepatocytes and HSCs) may arise from a common progenitor. Sca+ murine progenitor cells were found to coexpress markers of epithelial and mesenchymal lineages and to give rise, within few generations, to cells that segregate the lineage-specific markers into two distinct subpopulations. Notably, these progenitor cells, clonally derived, when transplanted in healthy livers, were found to generate epithelial and mesenchymal liver-specific derivatives (i.e. hepatocytes and HSCs) properly integrated in the liver architecture. These evidences suggest the existence of a 'bona fide' organ-specific meso-endodermal precursor cell, thus profoundly modifying current models of adult progenitor commitment believed, so far, to be lineage-restricted. Heterotopic transplantations, which confirm the dual differentiation potentiality of those cells, indicates as tissue local cues are necessary to drive a full hepatic differentiation. These data provide first evidences for an adult stem/precursor cell capable to differentiate in both parenchymal and non-parenchymal organ-specific components and candidate the liver as the instructive site for the reservoir compartment of HSC precursors as yet non-localized in the adult.
Figures
References
-
- Dellavalle A, Sampaolesi M, Tonlorenzi R, Tagliafico E, Sacchetti B, Perani L, et al. Pericytes of human skeletal muscle are myogenic precursors distinct from satellite cells. Nat Cell Biol. 2007;9:255–267. - PubMed
-
- Tanaka EM, Ferretti P. Considering the evolution of regeneration in the central nervous system. Nat Rev Neurosci. 2009;10:713–723. - PubMed
-
- Sell S. Liver stem cells. Mod Pathol. 1994;7:105–112. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
