

Actin cross-linking protein palladin and spermatogenesis
- PMID: 23687615
- PMCID: PMC3644046
- DOI: 10.4161/spmg.23473
Actin cross-linking protein palladin and spermatogenesis
Abstract
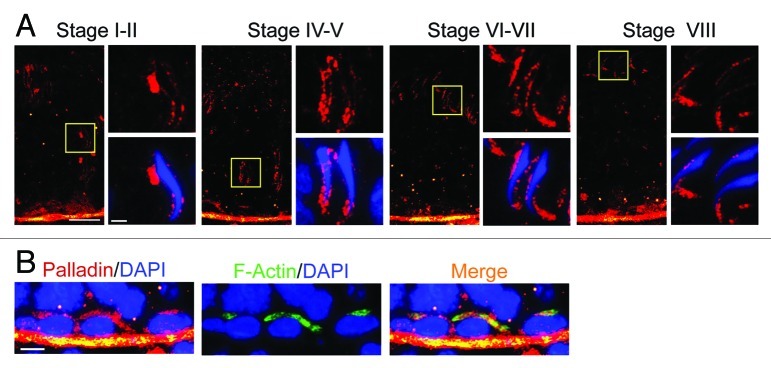
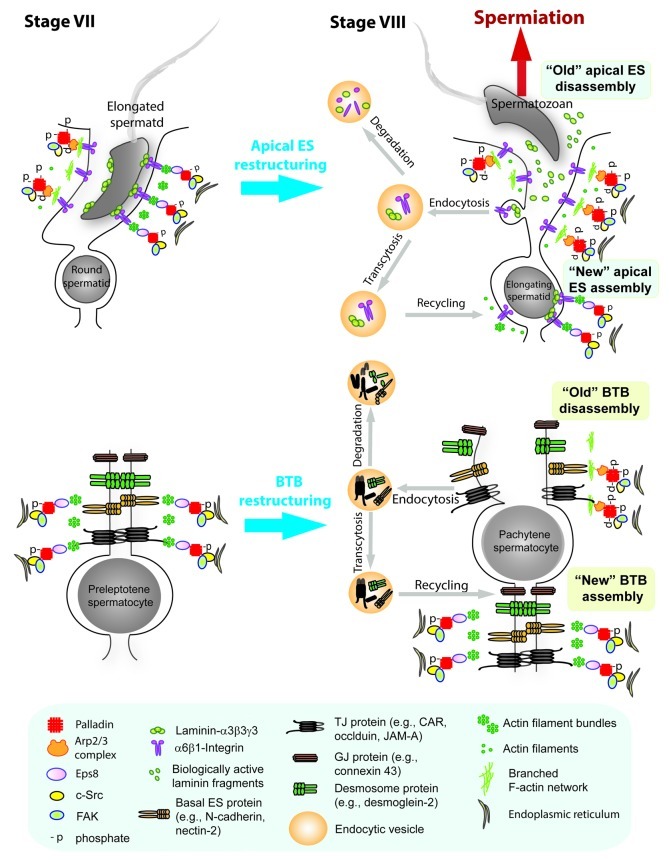
In the seminiferous epithelium of the mammalian testis, the most distinctive ultrastructure is the extensive bundles of actin filaments that lie near the Sertoli-spermatid interface and the Sertoli-Sertoli cell interface known as the apical ectoplasmic specialization (apical ES) and the basal ES, respectively. These actin filament bundles not only confer strong adhesion at these sites, they are uniquely found in the testis. Recent studies have shown that ES also confers spermatid and Sertoli cell polarity in the seminiferous epithelium during the epithelial cycle. While these junctions were first described in the 1970s, there are few functional studies in the literature to examine the regulation of these actin filament bundles. It is conceivable that these actin filament bundles at the ES undergo extensive re-organization to accommodate changes in location of developing spermatids during spermiogenesis as spermatids are transported across the seminiferous epithelium. Additionally, these actin filaments are rapidly reorganized during BTB restructuring to accommodate the transit of preleptotene spermatocytes across the barrier at stage VIII of the epithelial cycle. Thus, actin binding and regulatory proteins are likely involved in these events to confer changes in F-actin organization at these sites. Interestingly, there are no reports in the field to study these regulatory proteins until recently. Herein, we summarize some of the latest findings in the field regarding a novel actin cross-linker and actin-bundling protein called palladin. We also discuss in this opinion article the likely role of palladin in regulating actin filament bundles at the ES during spermatogenesis, highlighting the significant of palladin and how this protein is plausibly working in concert with other actin-binding/regulatory proteins and components of polarity proteins to regulate the cyclic events of actin organization and re-organization during the epithelial cycle of spermatogenesis. We also propose a hypothetic model by which palladin regulates ES restructuring during the epithelial cycle of spermatogenesis.
Keywords: F-actin; Sertoli cells; blood-testis barrier; ectoplasmic specialization; seminiferous epithelial cycle; spermatogenesis; testis; tight junction.
Figures

Similar articles
-
RAI14 (retinoic acid induced protein 14) is an F-actin regulator: Lesson from the testis.Spermatogenesis. 2013 Apr 1;3(2):e24824. doi: 10.4161/spmg.24824. Spermatogenesis. 2013. PMID: 23885305 Free PMC article.
-
Fascin - An actin binding and bundling protein in the testis and its role in ectoplasmic specialization dynamics.Spermatogenesis. 2015 Feb 23;5(1):e1002733. doi: 10.1080/21565562.2014.1002733. eCollection 2015 Jan-Apr. Spermatogenesis. 2015. PMID: 26413410 Free PMC article.
-
Actin-binding protein drebrin E is involved in junction dynamics during spermatogenesis.Spermatogenesis. 2011 Apr-Jun;1(2):123-136. doi: 10.4161/spmg.1.2.16393. Spermatogenesis. 2011. PMID: 22319661 Free PMC article.
-
Cell polarity proteins and spermatogenesis.Semin Cell Dev Biol. 2016 Nov;59:62-70. doi: 10.1016/j.semcdb.2016.06.008. Epub 2016 Jun 9. Semin Cell Dev Biol. 2016. PMID: 27292315 Free PMC article. Review.
-
Transport of germ cells across the seminiferous epithelium during spermatogenesis-the involvement of both actin- and microtubule-based cytoskeletons.Tissue Barriers. 2016 Nov 28;4(4):e1265042. doi: 10.1080/21688370.2016.1265042. eCollection 2016. Tissue Barriers. 2016. PMID: 28123928 Free PMC article. Review.
Cited by
-
RAI14 (retinoic acid induced protein 14) is an F-actin regulator: Lesson from the testis.Spermatogenesis. 2013 Apr 1;3(2):e24824. doi: 10.4161/spmg.24824. Spermatogenesis. 2013. PMID: 23885305 Free PMC article.
-
Actin binding proteins in blood-testis barrier function.Curr Opin Endocrinol Diabetes Obes. 2015 Jun;22(3):238-47. doi: 10.1097/MED.0000000000000155. Curr Opin Endocrinol Diabetes Obes. 2015. PMID: 25887390 Free PMC article. Review.
-
Toxicants target cell junctions in the testis: Insights from the indazole-carboxylic acid model.Spermatogenesis. 2015 Jan 21;4(2):e981485. doi: 10.4161/21565562.2014.981485. eCollection 2014 May-Aug. Spermatogenesis. 2015. PMID: 26413399 Free PMC article. Review.
-
The cell-cell junctions of mammalian testes: I. The adhering junctions of the seminiferous epithelium represent special differentiation structures.Cell Tissue Res. 2014 Sep;357(3):645-65. doi: 10.1007/s00441-014-1906-9. Epub 2014 Jun 8. Cell Tissue Res. 2014. PMID: 24907851 Free PMC article.
-
Focal adhesion kinase and actin regulatory/binding proteins that modulate F-actin organization at the tissue barrier: Lesson from the testis.Tissue Barriers. 2013 Apr 1;1(2):e24252. doi: 10.4161/tisb.24252. Tissue Barriers. 2013. PMID: 24665388 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources