

Thymic stromal lymphopoietin activity is increased in nasal polyps of patients with chronic rhinosinusitis
- PMID: 23688414
- PMCID: PMC3759596
- DOI: 10.1016/j.jaci.2013.04.005
Thymic stromal lymphopoietin activity is increased in nasal polyps of patients with chronic rhinosinusitis
Abstract
Background: Chronic rhinosinusitis with nasal polyps (CRSwNP) is associated with TH2-dominant inflammation. Thymic stromal lymphopoietin (TSLP) is a cytokine that triggers dendritic cell-mediated TH2 inflammatory responses and that enhances IL-1-dependent TH2 cytokine production in mast cells. Although increased TSLP mRNA levels have been found in nasal polyps (NPs), expression of TSLP protein and its function in patients with chronic rhinosinusitis (CRS) have not been fully explored.
Objectives: The objective of this study was to investigate the role of TSLP in patients with CRS.
Methods: We investigated the presence and stability of TSLP protein in NPs using ELISA and Western blotting and investigated the function of TSLP in nasal tissue extracts with a bioassay based on activation of human mast cells.
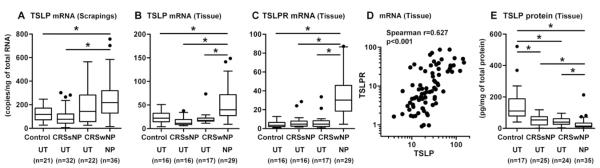
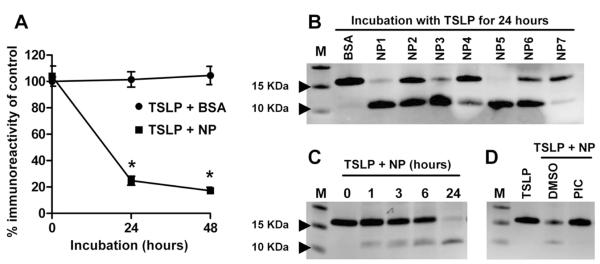
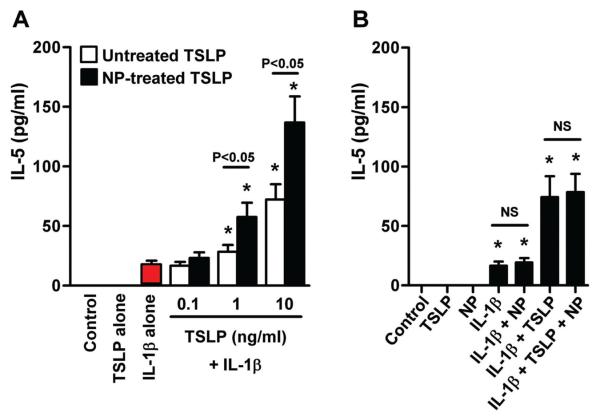
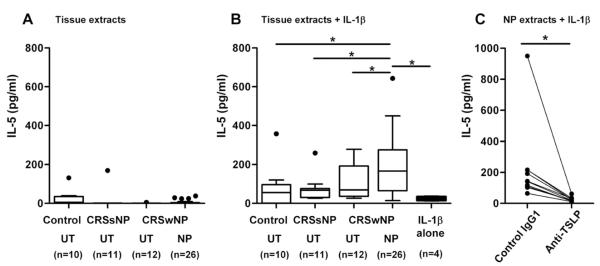
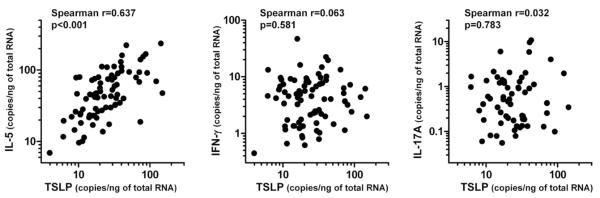
Results: Although TSLP mRNA levels were significantly increased in NP tissue from patients with CRSwNP compared with uncinate tissue from patients with CRS or control subjects, TSLP protein was significantly decreased in NP tissue, as detected by using the commercial ELISA kit. We found that recombinant TSLP was time-dependently degraded by NP extracts, and this degradation was completely inhibited by a protease inhibitor cocktail, suggesting that TSLP is sensitive to tissue proteases. Interestingly, NP extract-treated TSLP had higher activity in mast cells, although the amount of full-length TSLP was reduced up to 85%. NP extracts significantly enhanced IL-1β-dependent IL-5 production in mast cells compared with uncinate tissue homogenates, and responses were significantly inhibited by anti-TSLP, suggesting that NPs contain biologically relevant levels of TSLP activity.
Conclusion: TSLP and its metabolic products might play an important role in the inflammation seen in patients with CRSwNP.
Keywords: 2-ME; 2-Mercaptoethanol; CRS; CRSsNP; CRSwNP; Chronic rhinosinusitis; Chronic rhinosinusitis with nasal polyps; Chronic rhinosinusitis without nasal polyps; DC; Dendritic cell; Double-stranded RNA; IL-5; ILC2; IMDM; Iscove modified Dulbecco medium; Myeloid dendritic cell; NHBE; NMFF; NP; Nasal polyp; Normal human bronchial epithelial cell; Northwestern Medical Faculty Foundation; PIC; PNEC; Primary nasal epithelial cell; Protease inhibitor cocktail; SCF; Stem cell factor; T(H)2 cells; TLR; TSLP; TSLPR; Thymic stromal lymphopoietin; Thymic stromal lymphopoietin receptor; Toll-like receptor; Type 2 innate lymphoid cell; UT; Uncinate tissue; dsRNA; epithelial cells; mDC; mast cells; nasal polyps; proteases; thymic stromal lymphopoietin.
Copyright © 2013 American Academy of Allergy, Asthma & Immunology. Published by Mosby, Inc. All rights reserved.
Figures
References
-
- Bhattacharyya N. Incremental health care utilization and expenditures for chronic rhinosinusitis in the United States. Ann Otol Rhinol Laryngol. 2011;120:423–7. - PubMed
-
- Bachert C, Van Bruaene N, Toskala E, Zhang N, Olze H, Scadding G, et al. Important research questions in allergy and related diseases: 3-chronic rhinosinusitis and nasal polyposis - a GALEN study. Allergy. 2009;64:520–33. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
