

Secondary evolution of a self-incompatibility locus in the Brassicaceae genus Leavenworthia
- PMID: 23690750
- PMCID: PMC3653793
- DOI: 10.1371/journal.pbio.1001560
Secondary evolution of a self-incompatibility locus in the Brassicaceae genus Leavenworthia
Abstract
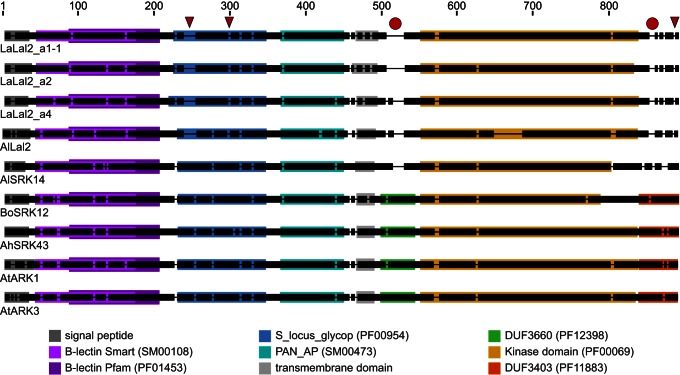
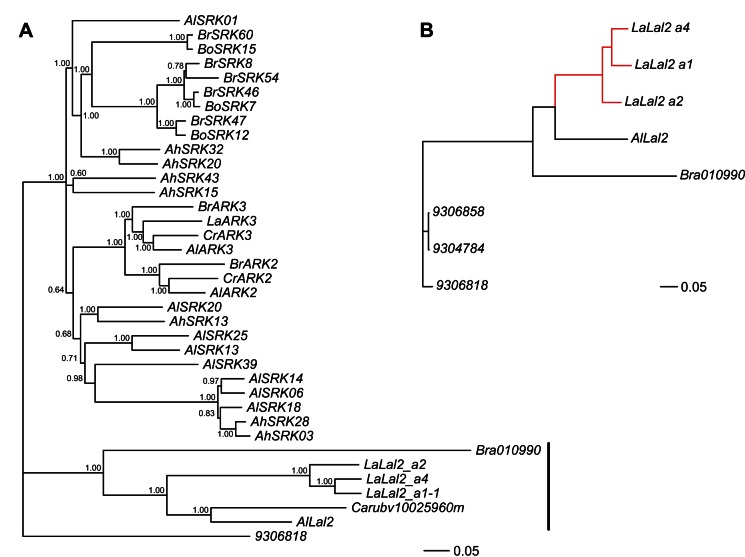
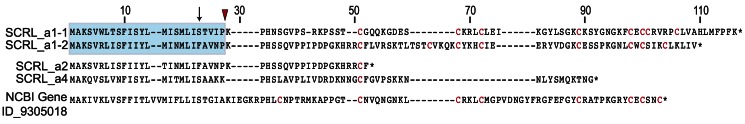
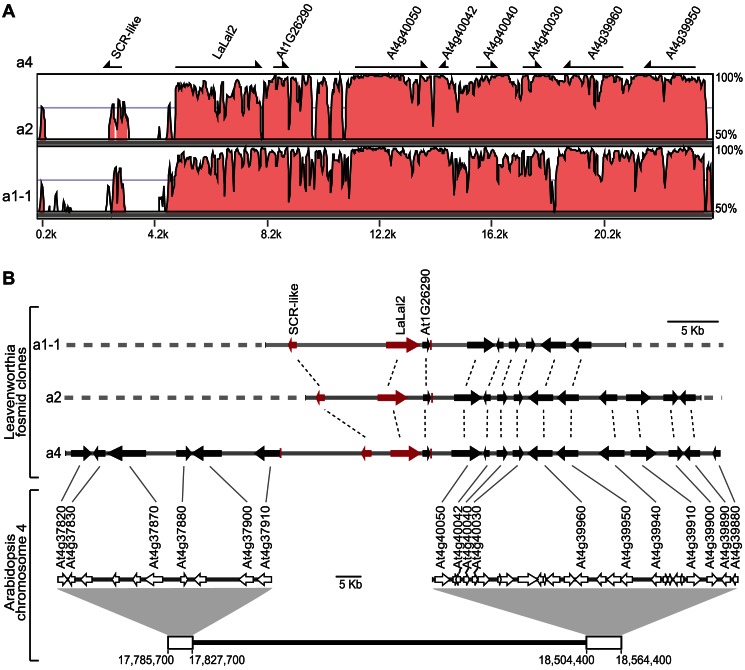
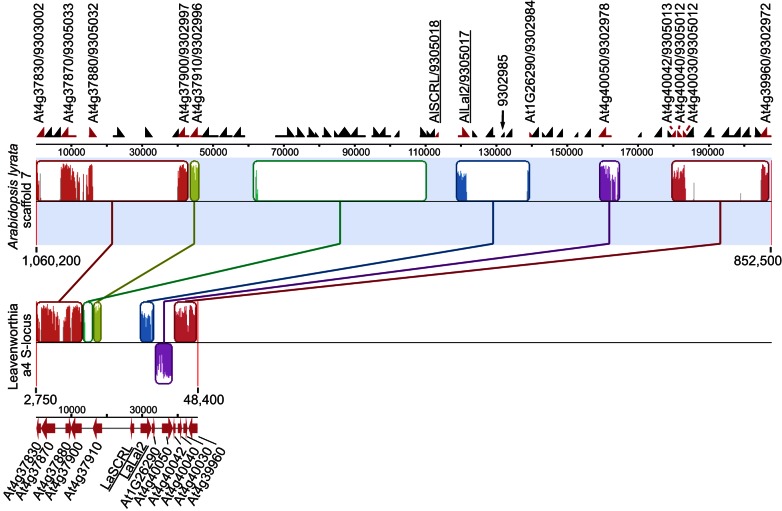
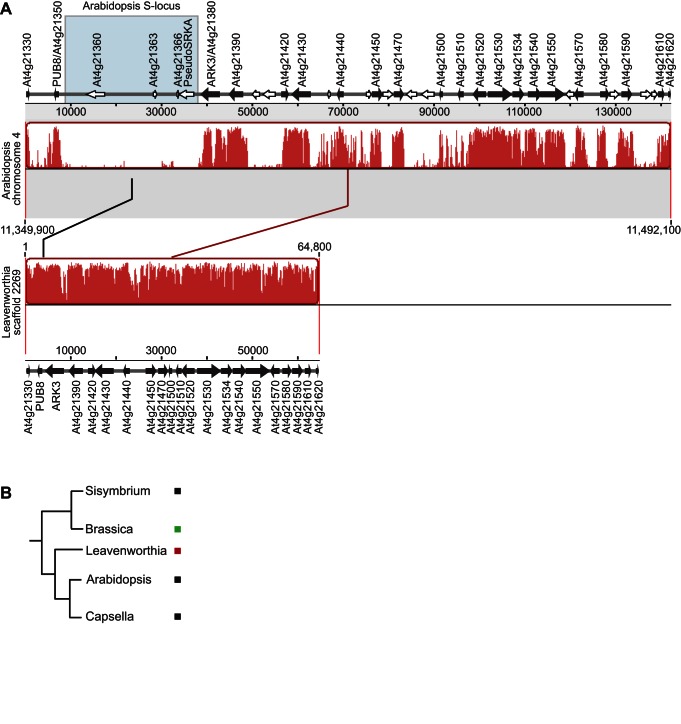
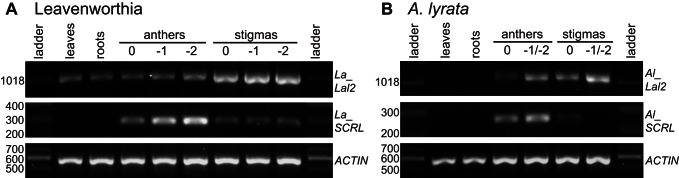
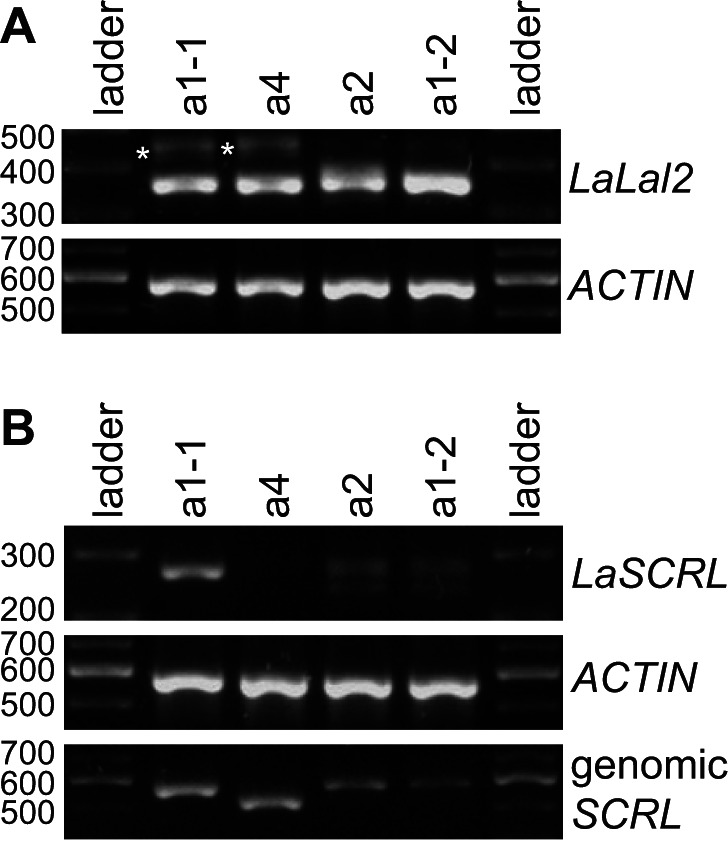
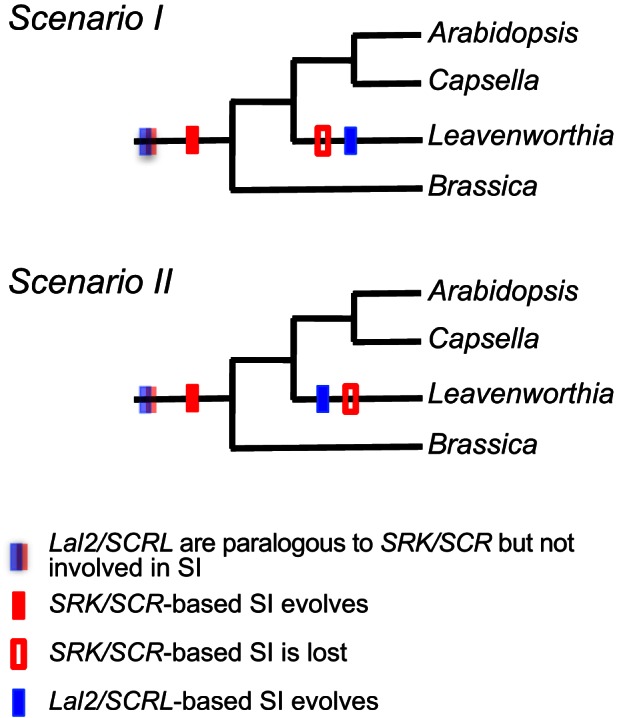
Self-incompatibility (SI) is the flowering plant reproductive system in which self pollen tube growth is inhibited, thereby preventing self-fertilization. SI has evolved independently in several different flowering plant lineages. In all Brassicaceae species in which the molecular basis of SI has been investigated in detail, the product of the S-locus receptor kinase (SRK) gene functions as receptor in the initial step of the self pollen-rejection pathway, while that of the S-locus cysteine-rich (SCR) gene functions as ligand. Here we examine the hypothesis that the S locus in the Brassicaceae genus Leavenworthia is paralogous with the S locus previously characterized in other members of the family. We also test the hypothesis that self-compatibility in this group is based on disruption of the pollen ligand-producing gene. Sequence analysis of the S-locus genes in Leavenworthia, phylogeny of S alleles, gene expression patterns, and comparative genomics analyses provide support for both hypotheses. Of special interest are two genes located in a non-S locus genomic region of Arabidopsis lyrata that exhibit domain structures, sequences, and phylogenetic histories similar to those of the S-locus genes in Leavenworthia, and that also share synteny with these genes. These A. lyrata genes resemble those comprising the A. lyrata S locus, but they do not function in self-recognition. Moreover, they appear to belong to a lineage that diverged from the ancestral Brassicaceae S-locus genes before allelic diversification at the S locus. We hypothesize that there has been neo-functionalization of these S-locus-like genes in the Leavenworthia lineage, resulting in evolution of a separate ligand-receptor system of SI. Our results also provide support for theoretical models that predict that the least constrained pathway to the evolution of self-compatibility is one involving loss of pollen gene function.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Whitehouse HLK (1950) Multiple-allelomorph incompatibility of pollen and style in the evolution of the Angiosperms. Ann Bot 14: 199–216.
-
- De Nettancourt D (1977) Incompatibility in Angiosperms. Berlin: Springer-Verlag. 230 p.
-
- Bateman AJ (1952) Self-incompatibility in Angiosperms. I. Theory. Heredity 6: 285–310.
-
- Franklin-Tong VE (2008) Self-incompatibility in flowering plants: evolution, diversity, and mechanisms. Springer. 341 p.
-
- Igic B, Bohs L, Kohn JR (2004) Historical inferences from the self-incompatibility locus. New Phytol 161: 97–105 doi:10.1046/j.1469-8137.2003.00952.x. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
