

Analysis of adult neurogenesis: evidence for a prominent "non-neurogenic" DCX-protein pool in rodent brain
- PMID: 23690918
- PMCID: PMC3653925
- DOI: 10.1371/journal.pone.0059269
Analysis of adult neurogenesis: evidence for a prominent "non-neurogenic" DCX-protein pool in rodent brain
Abstract
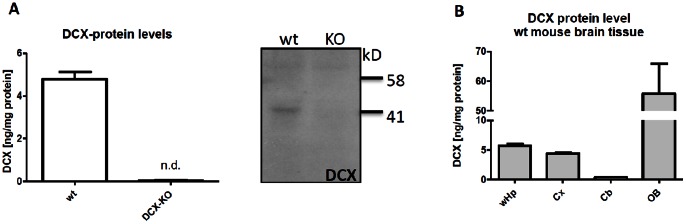
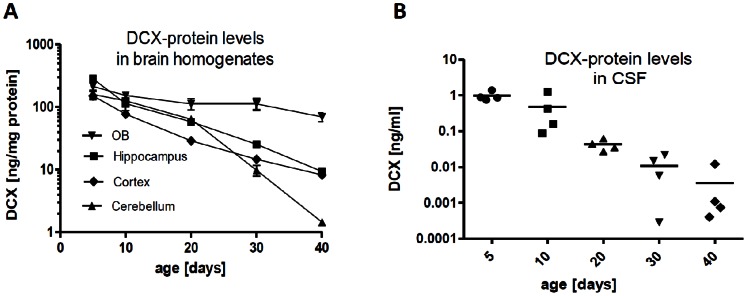
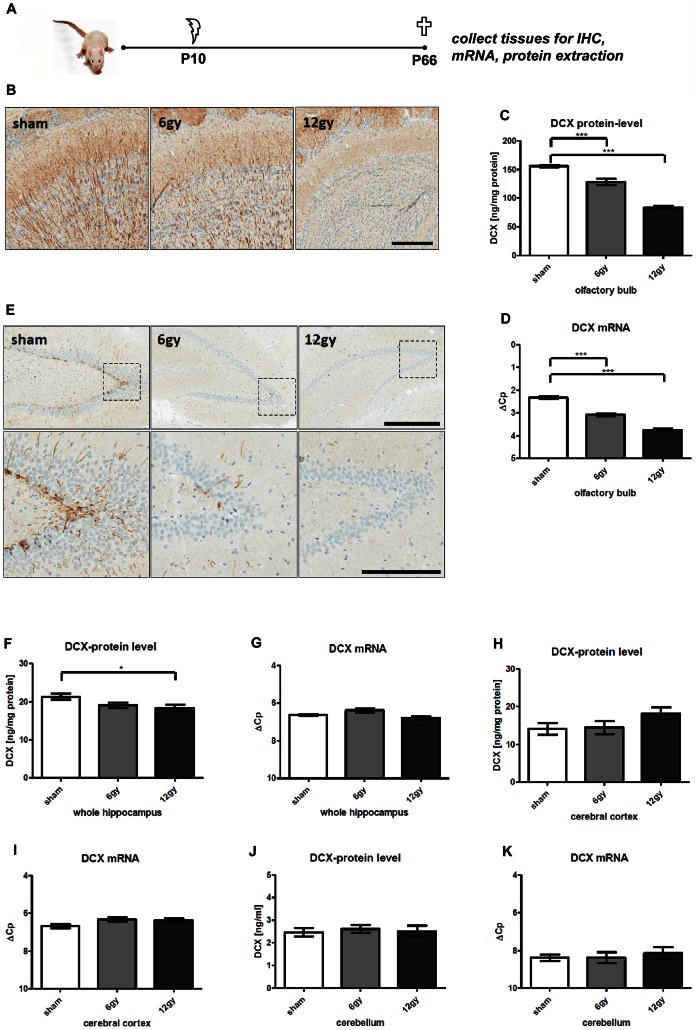
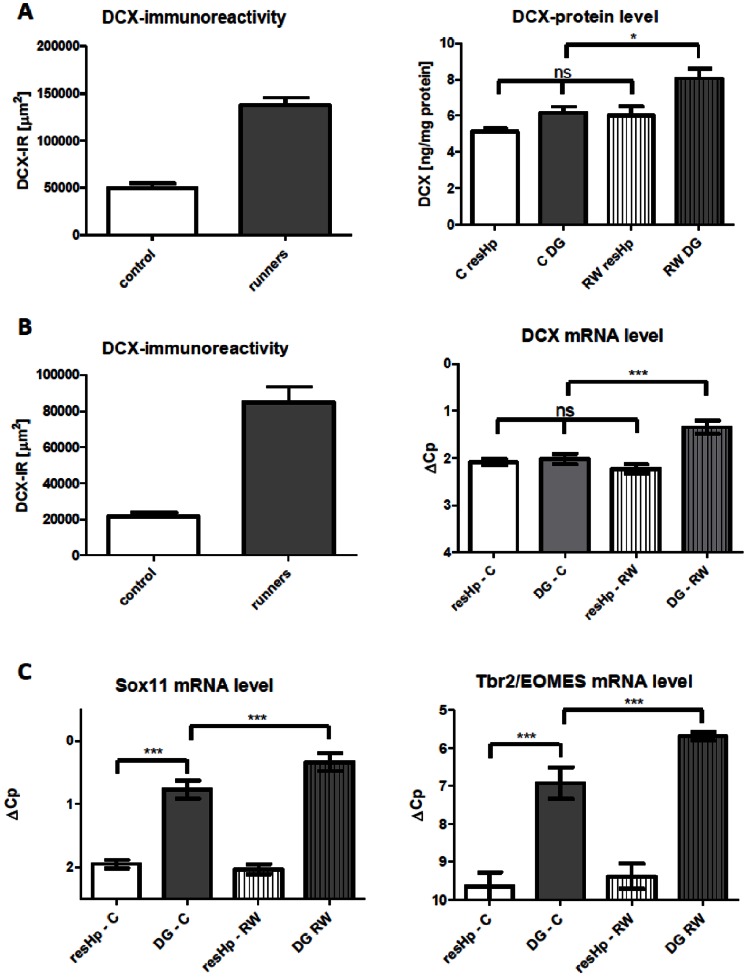
Here, we have developed a highly sensitive immunoassay for Dcx to characterize expression in brain and cerebrospinal fluid (CSF) of rodents. We demonstrate that Dcx is widely expressed during development in various brain regions and as well can be detected in cerebrospinal fluid of rats (up to 30 days postnatal). While Dcx protein level decline in adulthood and were detectable in neurogenic regions of the adult rodent brain, similar levels were also detectable in brain regions expected to bear no neurogenesis including the cerebral cortex and CA1/CA3 enriched hippocampus. We monitored DCX protein levels after paradigms to increase or severely decrease adult hippocampal neurogenesis, namely physical activity and cranial radiation, respectively. In both paradigms, Dcx protein- and mRNA-levels clearly reflected changes in neurogenesis in the hippocampus. However, basal Dcx-levels are unaffected in non-neurogenic regions (e.g. CA1/CA3 enriched hippocampus, cortex). These data suggest that there is a substantial "non-neurogenic" pool of Dcx- protein, whose regulation can be uncoupled from adult neurogenesis suggesting caution for the interpretation of such studies.
Conflict of interest statement
Figures
References
-
- Eriksson PS, Perfilieva E, Bjork-Eriksson T, Alborn AM, Nordborg C, et al. (1998) Neurogenesis in the adult human hippocampus. Nature medicine 4: 1313–1317. - PubMed
-
- Wolf SA, Melnik A, Kempermann G (2011) Physical exercise increases adult neurogenesis and telomerase activity, and improves behavioral deficits in a mouse model of schizophrenia. Brain, behavior, and immunity 25: 971–980. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
