

ZBTB33 binds unmethylated regions of the genome associated with actively expressed genes
- PMID: 23693142
- PMCID: PMC3663758
- DOI: 10.1186/1756-8935-6-13
ZBTB33 binds unmethylated regions of the genome associated with actively expressed genes
Abstract
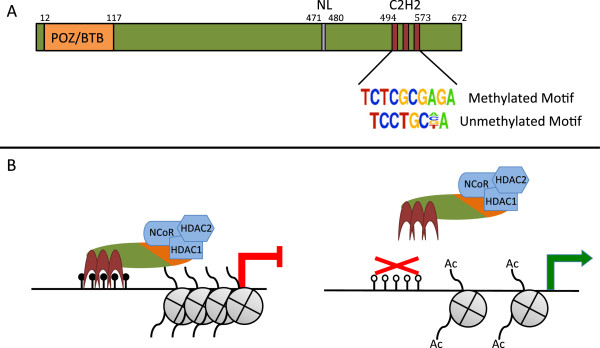
Background: DNA methylation and repressive histone modifications cooperate to silence promoters. One mechanism by which regions of methylated DNA could acquire repressive histone modifications is via methyl DNA-binding transcription factors. The zinc finger protein ZBTB33 (also known as Kaiso) has been shown in vitro to bind preferentially to methylated DNA and to interact with the SMRT/NCoR histone deacetylase complexes. We have performed bioinformatic analyses of Kaiso ChIP-seq and DNA methylation datasets to test a model whereby binding of Kaiso to methylated CpGs leads to loss of acetylated histones at target promoters.
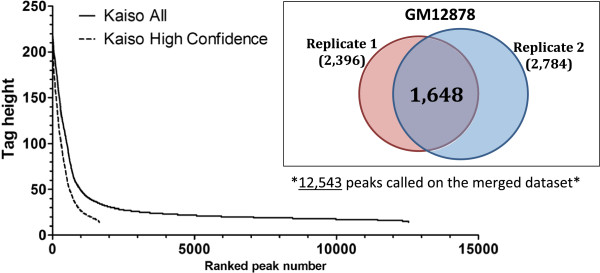
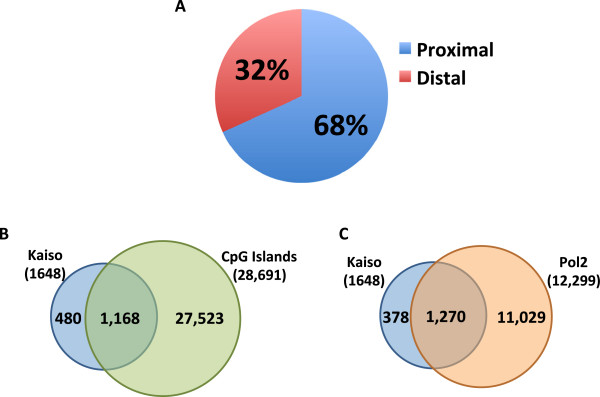
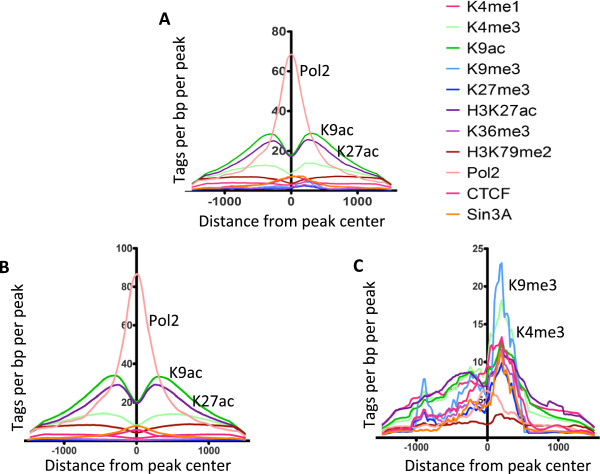
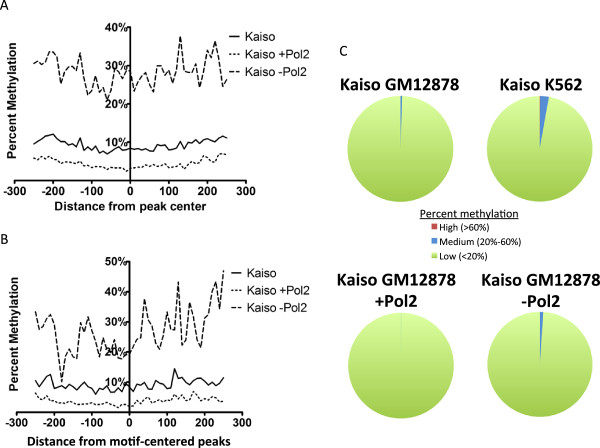
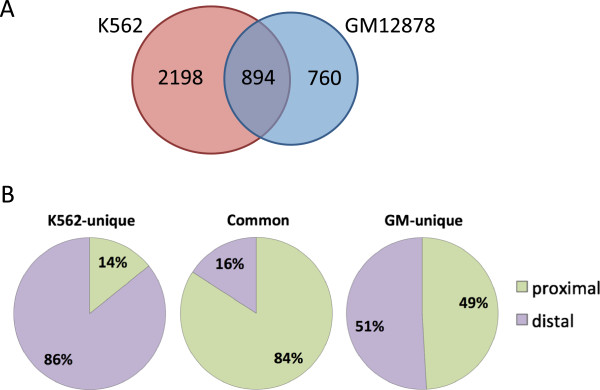
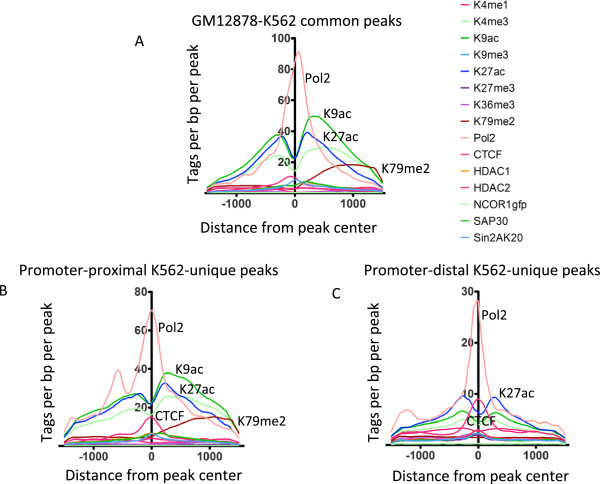
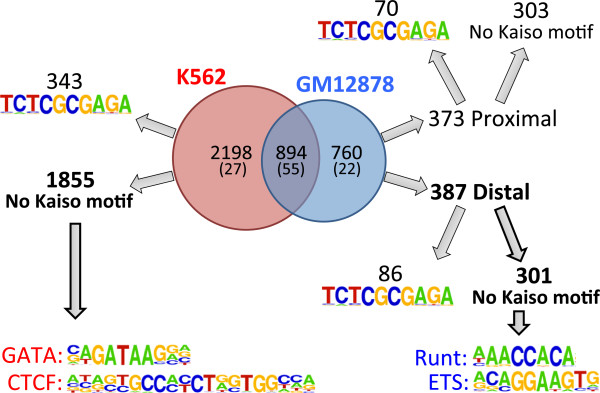
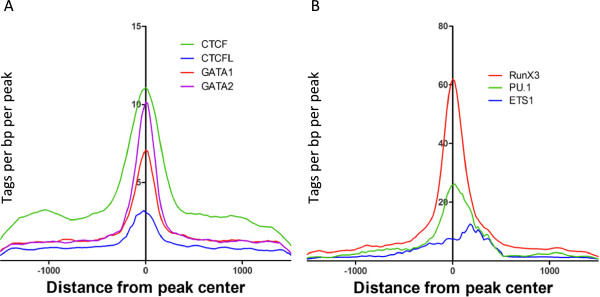
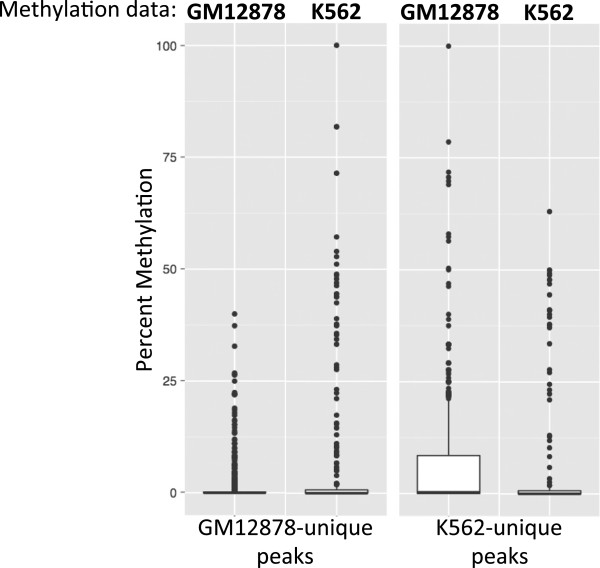
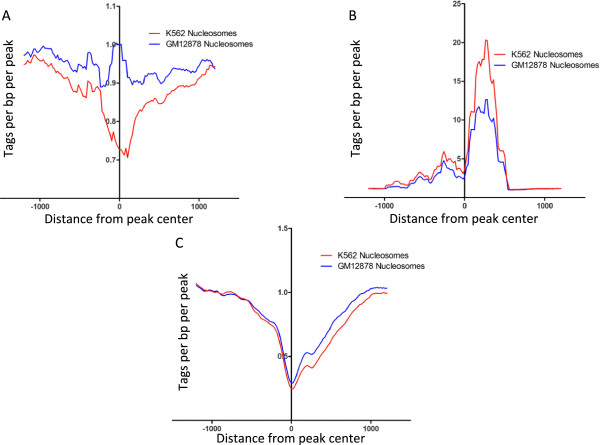
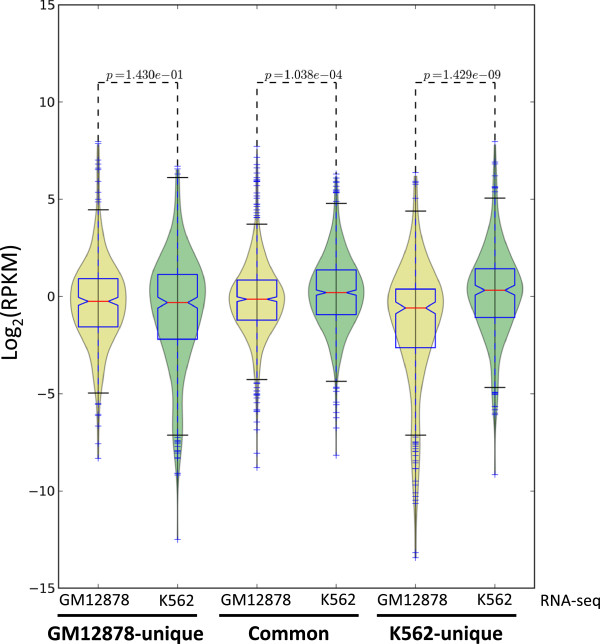
Results: Our results suggest that, contrary to expectations, Kaiso does not bind to methylated DNA in vivo but instead binds to highly active promoters that are marked with high levels of acetylated histones. In addition, our studies suggest that DNA methylation and nucleosome occupancy patterns restrict access of Kaiso to potential binding sites and influence cell type-specific binding.
Conclusions: We propose a new model for the genome-wide binding and function of Kaiso whereby Kaiso binds to unmethylated regulatory regions and contributes to the active state of target promoters.
Figures
References
-
- Brinkman AB, Gu H, Bartels SJJ, Zhang Y, Matarese F, Simmer F, Marks H, Bock C, Gnirke A, Meissner A, Stunnenberg HG. Sequential ChIP-bisulfite sequencing enables direct genome-scale investigation of chromatin and DNA methylation cross-talk. Genome Res. 2012;22:1128–1138. doi: 10.1101/gr.133728.111. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
