

Competition between model protocells driven by an encapsulated catalyst
- PMID: 23695631
- PMCID: PMC4041014
- DOI: 10.1038/nchem.1650
Competition between model protocells driven by an encapsulated catalyst
Erratum in
- Nat Chem. 2013 Jul;5(7):634
Abstract
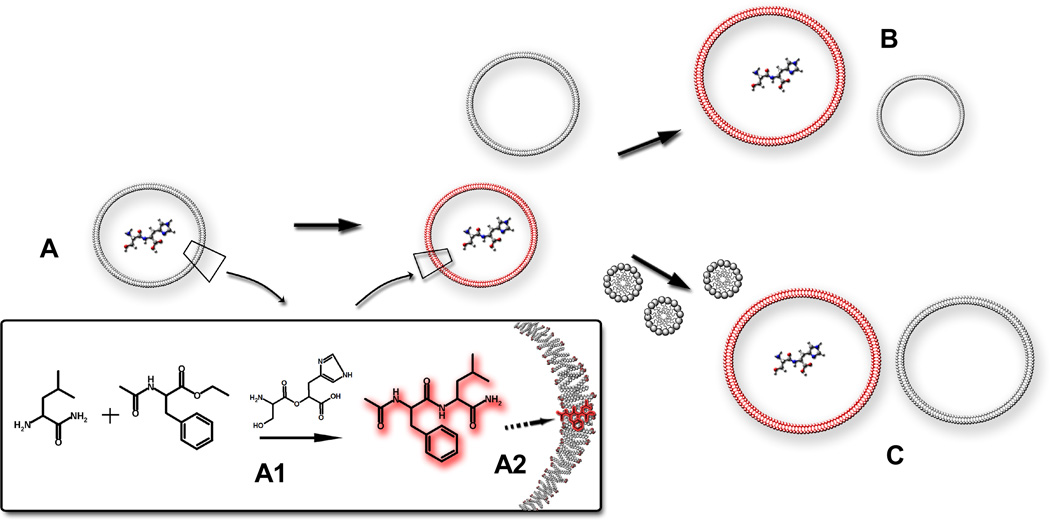
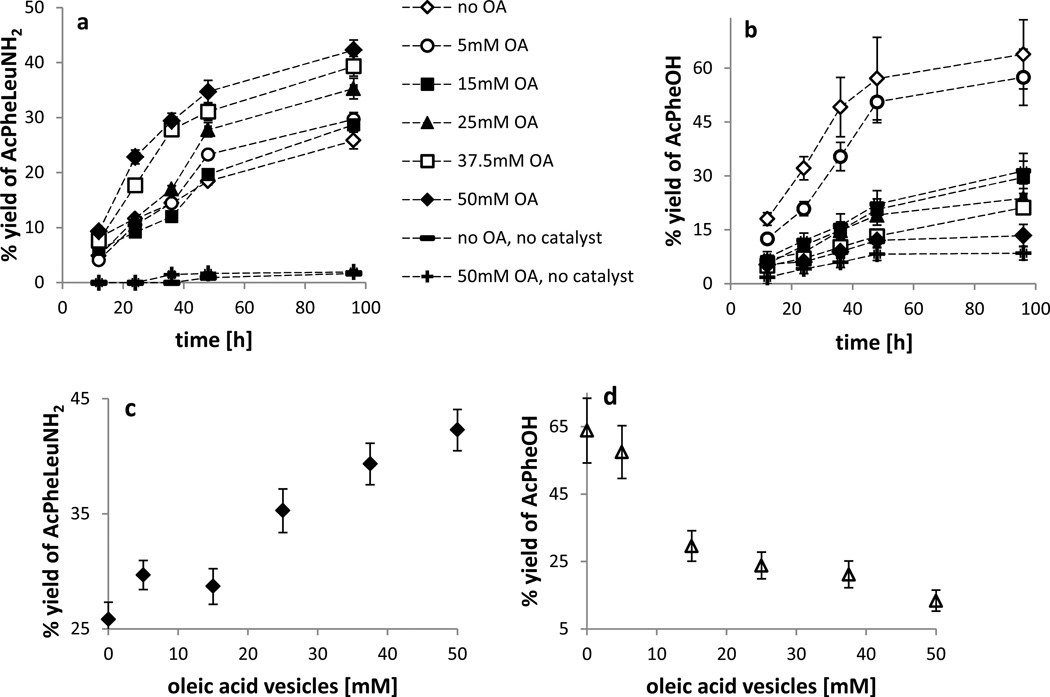
The advent of Darwinian evolution required the emergence of molecular mechanisms for the heritable variation of fitness. One model for such a system involves competing protocell populations, each consisting of a replicating genetic polymer within a replicating vesicle. In this model, each genetic polymer imparts a selective advantage to its protocell by, for example, coding for a catalyst that generates a useful metabolite. Here, we report a partial model of such nascent evolutionary traits in a system that consists of fatty-acid vesicles containing a dipeptide catalyst, which catalyses the formation of a second dipeptide. The newly formed dipeptide binds to vesicle membranes, which imparts enhanced affinity for fatty acids and thus promotes vesicle growth. The catalysed dipeptide synthesis proceeds with higher efficiency in vesicles than in free solution, which further enhances fitness. Our observations suggest that, in a replicating protocell with an RNA genome, ribozyme-catalysed peptide synthesis might have been sufficient to initiate Darwinian evolution.
Figures





References
-
- Monnard PA, Deamer DW. Membrane self-assembly processes: steps toward the first cellular life. The Anatomical record. 2002;268(3):196–207. - PubMed
-
- Szathmary E, Demeter L. Group selection of early replicators and the origin of life. J Theor Biol. 1987;128(4):463–486. - PubMed
-
- Szostak JW, Bartel DP, Luisi PL. Synthesizing life. Nature. 2001;409(6818):387–390. - PubMed
-
- Gebicki JM, Hicks M. Ufasomes are stable particles surrounded by unsaturated fatty acid membranes. Nature. 1973;243(5404):232–234. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
