

Lysophosphatidic acid mediates myeloid differentiation within the human bone marrow microenvironment
- PMID: 23696850
- PMCID: PMC3655943
- DOI: 10.1371/journal.pone.0063718
Lysophosphatidic acid mediates myeloid differentiation within the human bone marrow microenvironment
Abstract
Lysophosphatidic acid (LPA) is a pleiotropic phospholipid present in the blood and certain tissues at high concentrations; its diverse effects are mediated through differential, tissue specific expression of LPA receptors. Our goal was to determine if LPA exerts lineage-specific effects during normal human hematopoiesis. In vitro stimulation of CD34+ human hematopoietic progenitors by LPA induced myeloid differentiation but had no effect on lymphoid differentiation. LPA receptors were expressed at significantly higher levels on Common Myeloid Progenitors (CMP) than either multipotent Hematopoietic Stem/Progenitor Cells (HSPC) or Common Lymphoid Progenitors (CLP) suggesting that LPA acts on committed myeloid progenitors. Functional studies demonstrated that LPA enhanced migration, induced cell proliferation and reduced apoptosis of isolated CMP, but had no effect on either HSPC or CLP. Analysis of adult and fetal human bone marrow sections showed that PPAP2A, (the enzyme which degrades LPA) was highly expressed in the osteoblastic niche but not in the perivascular regions, whereas Autotaxin (the enzyme that synthesizes LPA) was expressed in perivascular regions of the marrow. We propose that a gradient of LPA with the highest levels in peri-sinusoidal regions and lowest near the endosteal zone, regulates the localization, proliferation and differentiation of myeloid progenitors within the bone marrow marrow.
Conflict of interest statement
Figures
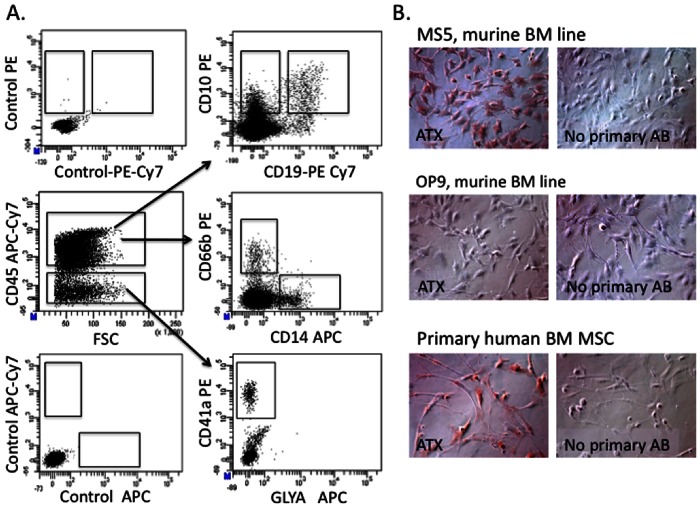
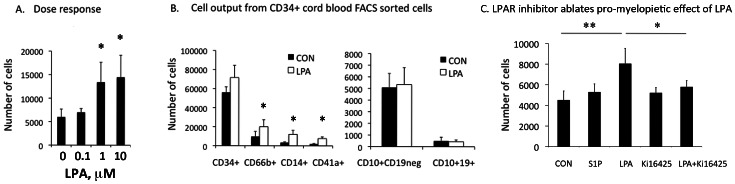
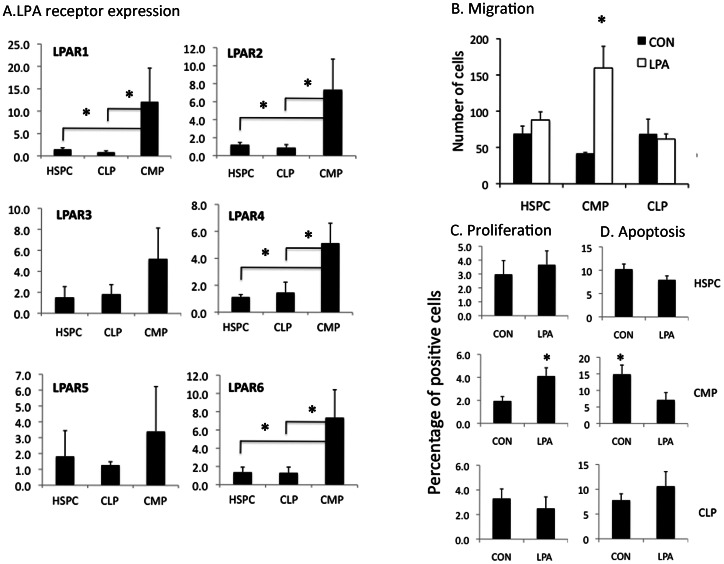
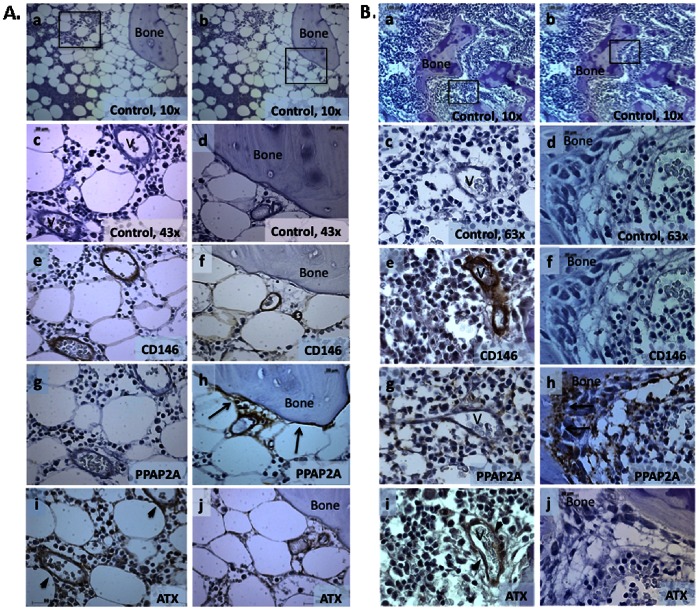
References
-
- Sano T, Baker D, Virag T, Wada A, Yatomi Y, et al. (2002) Multiple mechanisms linked to platelet activation result in lysophosphatidic acid and sphingosine 1-phosphate generation in blood. J Biol Chem 277: 21197–21206. - PubMed
-
- Tigyi G, Miledi R (1992) Lysophosphatidates bound to serum albumin activate membrane currents in Xenopus oocytes and neurite retraction in PC12 pheochromocytoma cells. J Biol Chem 267: 21360–21367. - PubMed
-
- Tokumura A, Majima E, Kariya Y, Tominaga K, Kogure K, et al. (2002) Identification of human plasma lysophospholipase D, a lysophosphatidic acid-producing enzyme, as autotaxin, a multifunctional phosphodiesterase. J Biol Chem 277: 39436–39442. - PubMed
-
- Blackburn J, Mansell JP (2012) The emerging role of lysophosphatidic acid (LPA) in skeletal biology. Bone 50: 756–762. - PubMed
-
- Choi JW, Herr DR, Noguchi K, Yung YC, Lee CW, et al. (2010) LPA receptors: subtypes and biological actions. Annu Rev Pharmacol Toxicol 50: 157–186. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
