

Molecular basis for the action of a dietary flavonoid revealed by the comprehensive identification of apigenin human targets
- PMID: 23697369
- PMCID: PMC3683737
- DOI: 10.1073/pnas.1303726110
Molecular basis for the action of a dietary flavonoid revealed by the comprehensive identification of apigenin human targets
Abstract
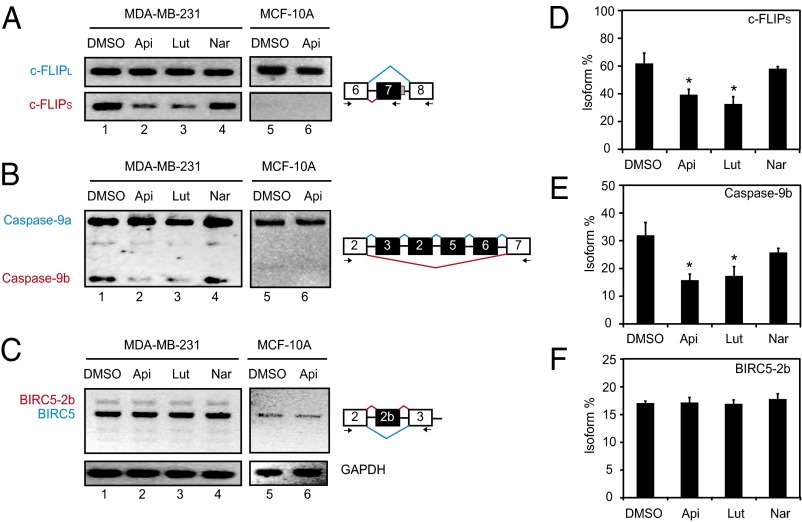
Flavonoids constitute the largest class of dietary phytochemicals, adding essential health value to our diet, and are emerging as key nutraceuticals. Cellular targets for dietary phytochemicals remain largely unknown, posing significant challenges for the regulation of dietary supplements and the understanding of how nutraceuticals provide health value. Here, we describe the identification of human cellular targets of apigenin, a flavonoid abundantly present in fruits and vegetables, using an innovative high-throughput approach that combines phage display with second generation sequencing. The 160 identified high-confidence candidate apigenin targets are significantly enriched in three main functional categories: GTPase activation, membrane transport, and mRNA metabolism/alternative splicing. This last category includes the heterogeneous nuclear ribonucleoprotein A2 (hnRNPA2), a factor involved in splicing regulation, mRNA stability, and mRNA transport. Apigenin binds to the C-terminal glycine-rich domain of hnRNPA2, preventing hnRNPA2 from forming homodimers, and therefore, it perturbs the alternative splicing of several human hnRNPA2 targets. Our results provide a framework to understand how dietary phytochemicals exert their actions by binding to many functionally diverse cellular targets. In turn, some of them may modulate the activity of a large number of downstream genes, which is exemplified here by the effects of apigenin on the alternative splicing activity of hnRNPA2. Hence, in contrast to small-molecule pharmaceuticals designed for defined target specificity, dietary phytochemicals affect a large number of cellular targets with varied affinities that, combined, result in their recognized health benefits.
Keywords: FRET; cancer; inflammation; nanosensor.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Heterogeneous nuclear ribonucleoprotein A2/B1 is a negative regulator of human breast cancer metastasis by maintaining the balance of multiple genes and pathways.EBioMedicine. 2020 Jan;51:102583. doi: 10.1016/j.ebiom.2019.11.044. Epub 2020 Jan 3. EBioMedicine. 2020. PMID: 31901866 Free PMC article.
-
Apigenin by targeting hnRNPA2 sensitizes triple-negative breast cancer spheroids to doxorubicin-induced apoptosis and regulates expression of ABCC4 and ABCG2 drug efflux transporters.Biochem Pharmacol. 2020 Dec;182:114259. doi: 10.1016/j.bcp.2020.114259. Epub 2020 Oct 2. Biochem Pharmacol. 2020. PMID: 33011162 Free PMC article.
-
Transcriptome reprogramming through alternative splicing triggered by apigenin drives cell death in triple-negative breast cancer.Cell Death Dis. 2023 Dec 13;14(12):824. doi: 10.1038/s41419-023-06342-6. Cell Death Dis. 2023. PMID: 38092740 Free PMC article.
-
hnRNP A1: the Swiss army knife of gene expression.Int J Mol Sci. 2013 Sep 16;14(9):18999-9024. doi: 10.3390/ijms140918999. Int J Mol Sci. 2013. PMID: 24065100 Free PMC article. Review.
-
Heterogeneous nuclear ribonucleoprotein particle A/B proteins and the control of alternative splicing of the mammalian heterogeneous nuclear ribonucleoprotein particle A1 pre-mRNA.Prog Mol Subcell Biol. 2003;31:59-88. doi: 10.1007/978-3-662-09728-1_3. Prog Mol Subcell Biol. 2003. PMID: 12494763 Review. No abstract available.
Cited by
-
Dietary apigenin reduces LPS-induced expression of miR-155 restoring immune balance during inflammation.Mol Nutr Food Res. 2015 Apr;59(4):763-72. doi: 10.1002/mnfr.201400705. Epub 2015 Feb 23. Mol Nutr Food Res. 2015. PMID: 25641956 Free PMC article.
-
Bioengineering of Genetically Encoded Gene Promoter Repressed by the Flavonoid Apigenin for Constructing Intracellular Sensor for Molecular Events.Biosensors (Basel). 2021 Apr 28;11(5):137. doi: 10.3390/bios11050137. Biosensors (Basel). 2021. PMID: 33924783 Free PMC article.
-
Antioxidants in Translational Medicine.Antioxid Redox Signal. 2015 Nov 10;23(14):1130-43. doi: 10.1089/ars.2015.6393. Epub 2015 Aug 31. Antioxid Redox Signal. 2015. PMID: 26154592 Free PMC article. Review.
-
The Antioxidative Effects of Flavones in Hypertensive Disease.Biomedicines. 2023 Oct 24;11(11):2877. doi: 10.3390/biomedicines11112877. Biomedicines. 2023. PMID: 38001878 Free PMC article. Review.
-
IKKα Induces Epithelial-Mesenchymal Changes in Mouse Skin Carcinoma Cells That Can Be Partially Reversed by Apigenin.Int J Mol Sci. 2022 Jan 25;23(3):1375. doi: 10.3390/ijms23031375. Int J Mol Sci. 2022. PMID: 35163299 Free PMC article.
References
-
- Higdon J. An Evidence-Based Approach to Dietary Phytochemicals. New York: Thieme; 2007.
-
- Verpoorte R. Pharmacognosy in the new millennium: Leadfinding and biotechnology. J Pharm Pharmacol. 2000;52(3):253–262. - PubMed
-
- Clifford M, Brown JE. In: Flavonoids: Chemistry, Biochemistry and Applications. Andersen OM, Markham KR, editors. Boca Raton, FL: Taylor and Francis Group; 2006. pp. 320–370.
-
- Crozier A, Jaganath IB, Clifford MN. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat Prod Rep. 2009;26(8):1001–1043. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
