

Innate lymphoid cells regulate CD4+ T-cell responses to intestinal commensal bacteria
- PMID: 23698371
- PMCID: PMC3699860
- DOI: 10.1038/nature12240
Innate lymphoid cells regulate CD4+ T-cell responses to intestinal commensal bacteria
Abstract
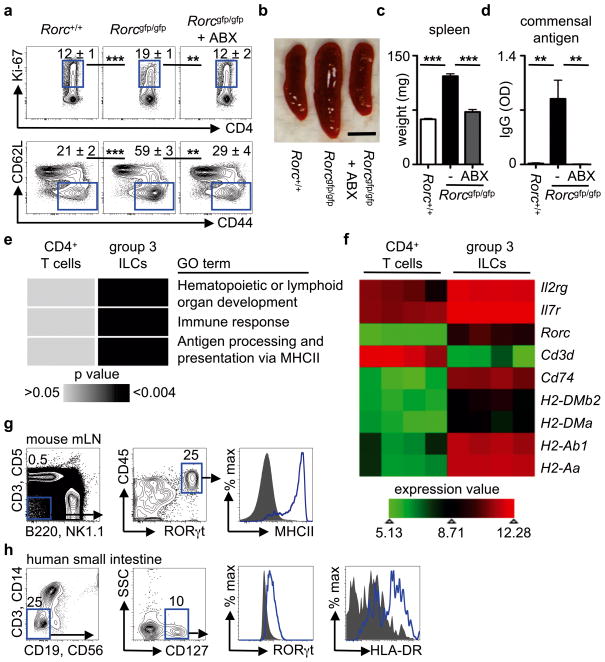
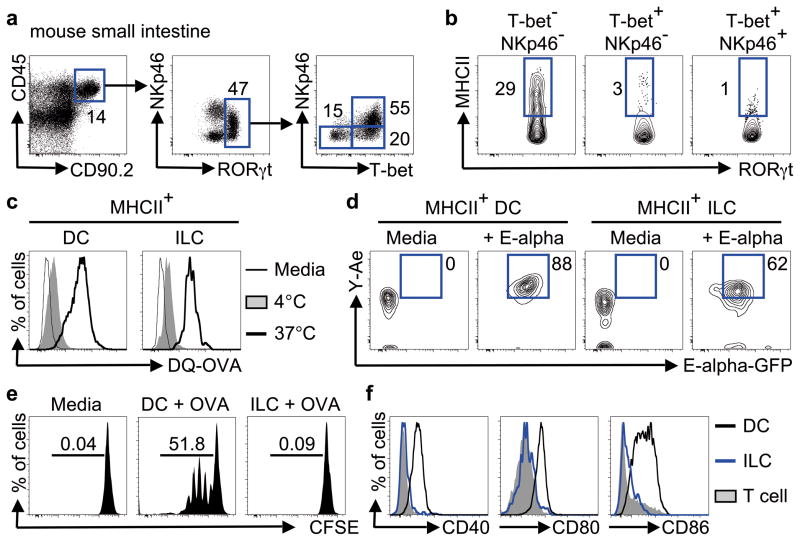
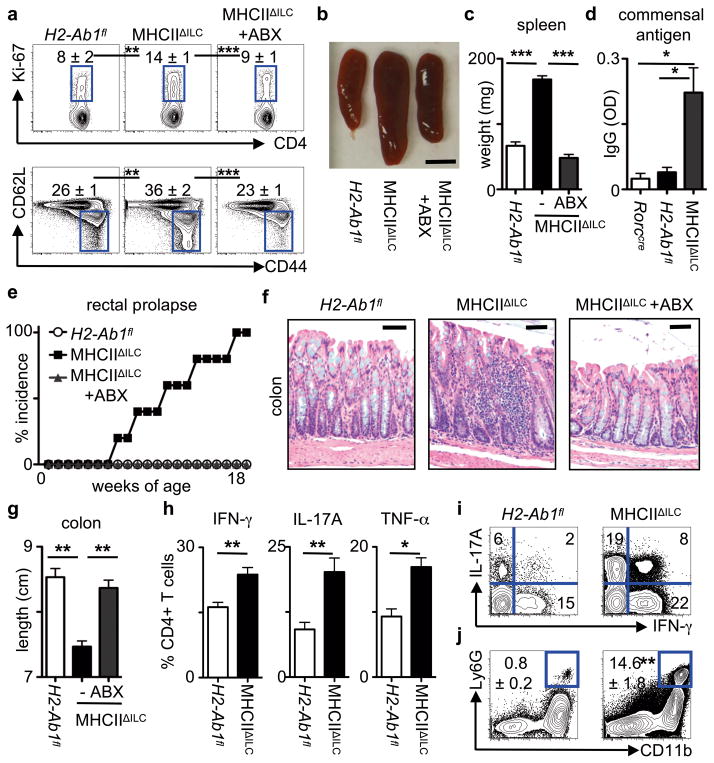
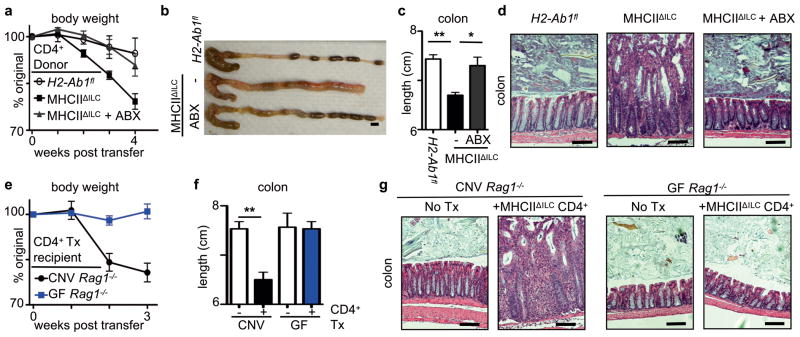
Innate lymphoid cells (ILCs) are a recently characterized family of immune cells that have critical roles in cytokine-mediated regulation of intestinal epithelial cell barrier integrity. Alterations in ILC responses are associated with multiple chronic human diseases, including inflammatory bowel disease, implicating a role for ILCs in disease pathogenesis. Owing to an inability to target ILCs selectively, experimental studies assessing ILC function have predominantly used mice lacking adaptive immune cells. However, in lymphocyte-sufficient hosts ILCs are vastly outnumbered by CD4(+) T cells, which express similar profiles of effector cytokines. Therefore, the function of ILCs in the presence of adaptive immunity and their potential to influence adaptive immune cell responses remain unknown. To test this, we used genetic or antibody-mediated depletion strategies to target murine ILCs in the presence of an adaptive immune system. We show that loss of retinoic-acid-receptor-related orphan receptor-γt-positive (RORγt(+)) ILCs was associated with dysregulated adaptive immune cell responses against commensal bacteria and low-grade systemic inflammation. Remarkably, ILC-mediated regulation of adaptive immune cells occurred independently of interleukin (IL)-17A, IL-22 or IL-23. Genome-wide transcriptional profiling and functional analyses revealed that RORγt(+) ILCs express major histocompatibility complex class II (MHCII) and can process and present antigen. However, rather than inducing T-cell proliferation, ILCs acted to limit commensal bacteria-specific CD4(+) T-cell responses. Consistent with this, selective deletion of MHCII in murine RORγt(+) ILCs resulted in dysregulated commensal bacteria-dependent CD4(+) T-cell responses that promoted spontaneous intestinal inflammation. These data identify that ILCs maintain intestinal homeostasis through MHCII-dependent interactions with CD4(+) T cells that limit pathological adaptive immune cell responses to commensal bacteria.
Figures
Comment in
-
Immunology: An innate regulatory cell.Nature. 2013 Jun 6;498(7452):42-3. doi: 10.1038/498042a. Nature. 2013. PMID: 23739420 No abstract available.
-
Mucosal immunology: ILCs broker peace deals in the gut.Nat Rev Immunol. 2013 Jul;13(7):473. doi: 10.1038/nri3480. Epub 2013 Jun 7. Nat Rev Immunol. 2013. PMID: 23743477 No abstract available.
References
-
- Spits H, Cupedo T. Innate lymphoid cells: emerging insights in development, lineage relationships, and function. Annu Rev Immunol. 2012;30:647–675. - PubMed
-
- Sawa S, et al. RORgammat(+) innate lymphoid cells regulate intestinal homeostasis by integrating negative signals from the symbiotic microbiota. Nat Immunol. 2011;12:320–326. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- AI061570/AI/NIAID NIH HHS/United States
- T32-AI055428/AI/NIAID NIH HHS/United States
- AI102942/AI/NIAID NIH HHS/United States
- DK071176/DK/NIDDK NIH HHS/United States
- AI074878/AI/NIAID NIH HHS/United States
- AI087990/AI/NIAID NIH HHS/United States
- P30DK50306/DK/NIDDK NIH HHS/United States
- 2-P30 CA016520/CA/NCI NIH HHS/United States
- R01 AI102942/AI/NIAID NIH HHS/United States
- R21 AI087990/AI/NIAID NIH HHS/United States
- R21 AI083480/AI/NIAID NIH HHS/United States
- U01 AI095608/AI/NIAID NIH HHS/United States
- R01 AI061570/AI/NIAID NIH HHS/United States
- AI095776/AI/NIAID NIH HHS/United States
- R01 AI074878/AI/NIAID NIH HHS/United States
- T32 AI055428/AI/NIAID NIH HHS/United States
- R01 AI095466/AI/NIAID NIH HHS/United States
- P30 DK050306/DK/NIDDK NIH HHS/United States
- T32 AI007532/AI/NIAID NIH HHS/United States
- DP5OD012116/OD/NIH HHS/United States
- DP5 OD012116/OD/NIH HHS/United States
- AI095608/AI/NIAID NIH HHS/United States
- P01 DK071176/DK/NIDDK NIH HHS/United States
- P30 CA016520/CA/NCI NIH HHS/United States
- R01 AI097333/AI/NIAID NIH HHS/United States
- AI095466/AI/NIAID NIH HHS/United States
- U01 AI095776/AI/NIAID NIH HHS/United States
- AI097333/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
