

F-actin asymmetry and the endoplasmic reticulum-associated TCC-1 protein contribute to stereotypic spindle movements in the Caenorhabditis elegans embryo
- PMID: 23699393
- PMCID: PMC3708726
- DOI: 10.1091/mbc.E13-02-0076
F-actin asymmetry and the endoplasmic reticulum-associated TCC-1 protein contribute to stereotypic spindle movements in the Caenorhabditis elegans embryo
Abstract
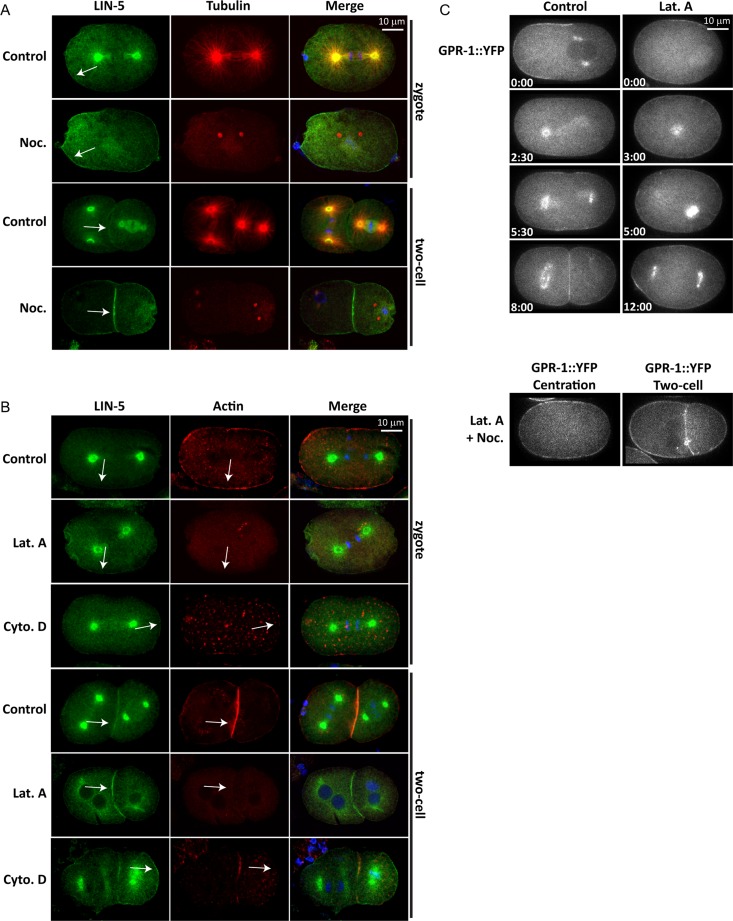
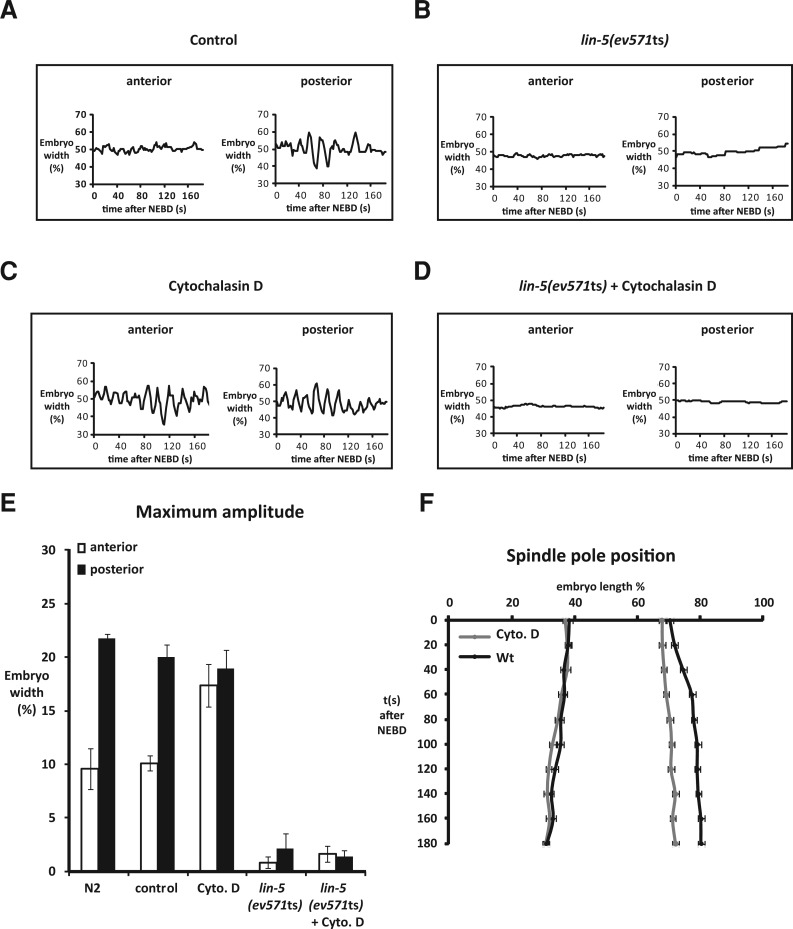
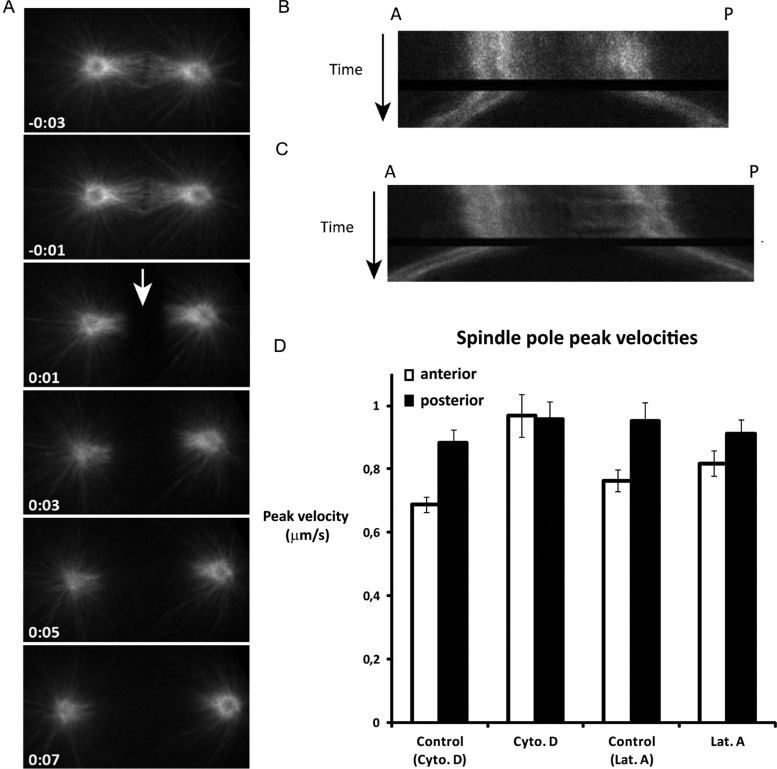
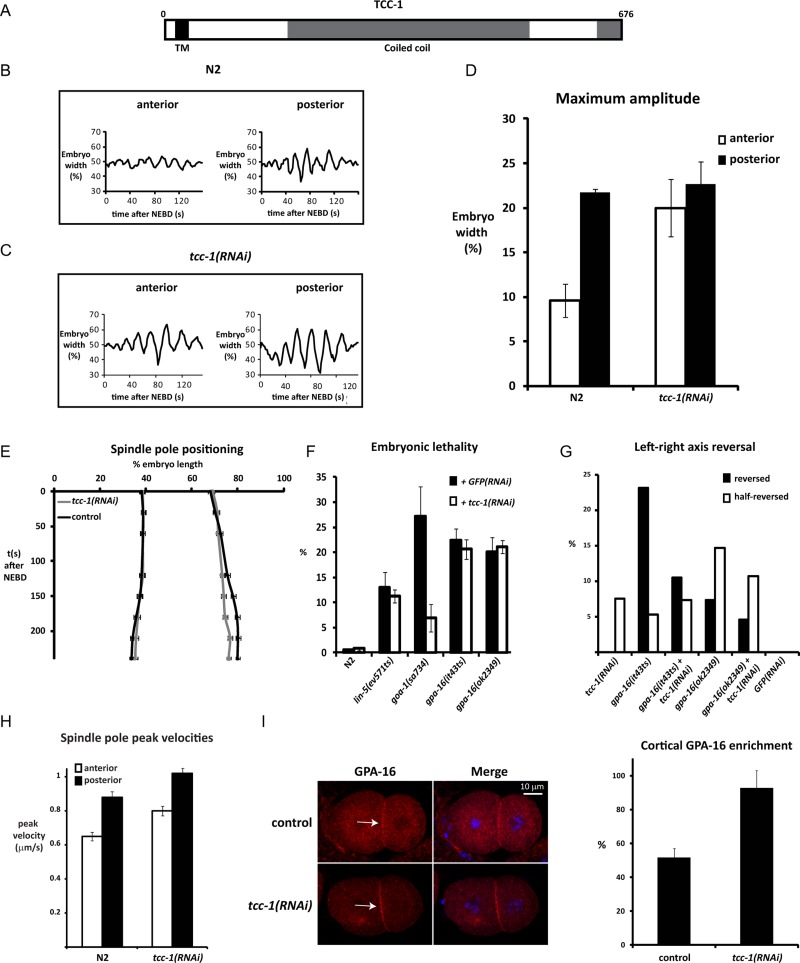
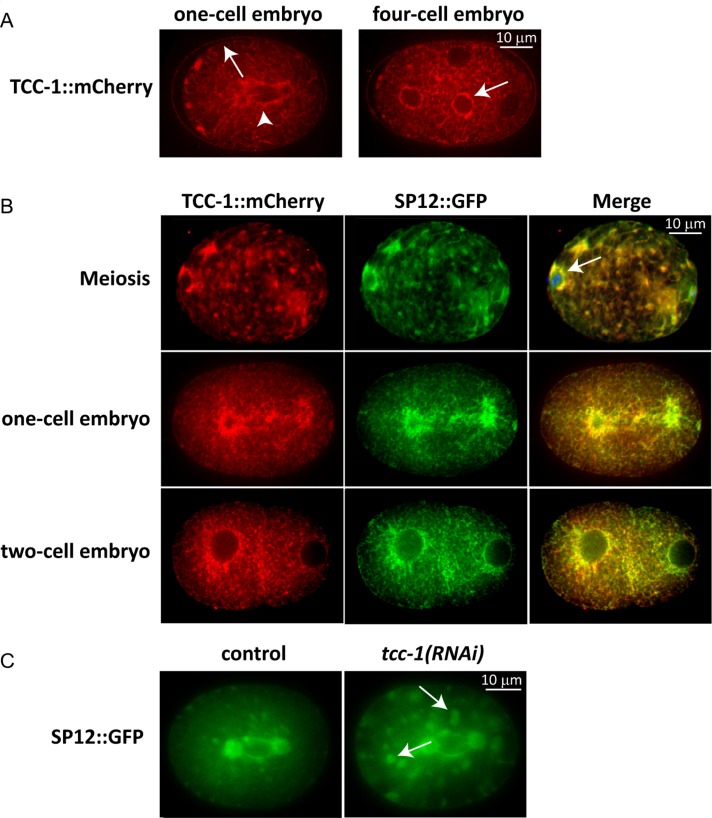
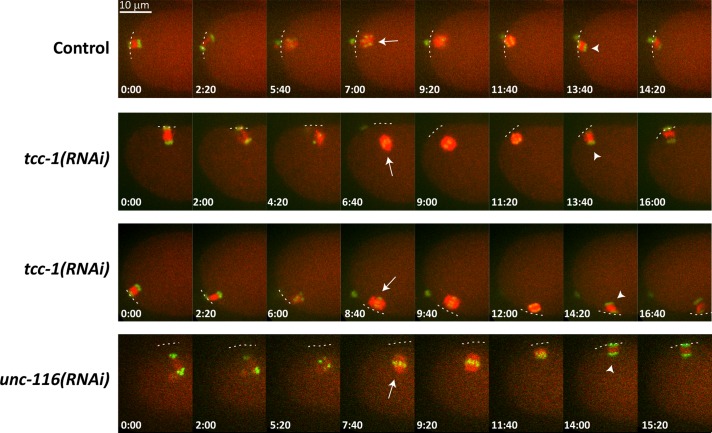
The microtubule spindle apparatus dictates the plane of cell cleavage in animal cells. During development, dividing cells control the position of the spindle to determine the size, location, and fate of daughter cells. Spindle positioning depends on pulling forces that act between the cell periphery and astral microtubules. This involves dynein recruitment to the cell cortex by a heterotrimeric G-protein α subunit in complex with a TPR-GoLoco motif protein (GPR-1/2, Pins, LGN) and coiled-coil protein (LIN-5, Mud, NuMA). In this study, we searched for additional factors that contribute to spindle positioning in the one-cell Caenorhabditis elegans embryo. We show that cortical actin is not needed for Gα-GPR-LIN-5 localization and pulling force generation. Instead, actin accumulation in the anterior actually reduces pulling forces, possibly by increasing cortical rigidity. Examining membrane-associated proteins that copurified with GOA-1 Gα, we found that the transmembrane and coiled-coil domain protein 1 (TCC-1) contributes to proper spindle movements. TCC-1 localizes to the endoplasmic reticulum membrane and interacts with UNC-116 kinesin-1 heavy chain in yeast two-hybrid assays. RNA interference of tcc-1 and unc-116 causes similar defects in meiotic spindle positioning, supporting the concept of TCC-1 acting with kinesin-1 in vivo. These results emphasize the contribution of membrane-associated and cortical proteins other than Gα-GPR-LIN-5 in balancing the pulling forces that position the spindle during asymmetric cell division.
Figures
Similar articles
-
Multisite Phosphorylation of NuMA-Related LIN-5 Controls Mitotic Spindle Positioning in C. elegans.PLoS Genet. 2016 Oct 6;12(10):e1006291. doi: 10.1371/journal.pgen.1006291. eCollection 2016 Oct. PLoS Genet. 2016. PMID: 27711157 Free PMC article.
-
aPKC phosphorylates NuMA-related LIN-5 to position the mitotic spindle during asymmetric division.Nat Cell Biol. 2011 Aug 21;13(9):1132-8. doi: 10.1038/ncb2315. Nat Cell Biol. 2011. PMID: 21857670
-
Mitotic Spindle Positioning in the EMS Cell of Caenorhabditis elegans Requires LET-99 and LIN-5/NuMA.Genetics. 2016 Nov;204(3):1177-1189. doi: 10.1534/genetics.116.192831. Epub 2016 Sep 26. Genetics. 2016. PMID: 27672093 Free PMC article.
-
Mechanisms of spindle positioning: cortical force generators in the limelight.Curr Opin Cell Biol. 2013 Dec;25(6):741-8. doi: 10.1016/j.ceb.2013.07.008. Epub 2013 Aug 16. Curr Opin Cell Biol. 2013. PMID: 23958212 Review.
-
[Mechanisms of cell division: lessons from a nematode].Med Sci (Paris). 2003 Jun-Jul;19(6-7):735-42. doi: 10.1051/medsci/20031967735. Med Sci (Paris). 2003. PMID: 12942445 Review. French.
Cited by
-
Num1 versus NuMA: insights from two functionally homologous proteins.Biophys Rev. 2018 Dec;10(6):1631-1636. doi: 10.1007/s12551-018-0472-x. Epub 2018 Nov 6. Biophys Rev. 2018. PMID: 30402673 Free PMC article. Review.
-
Tumor suppressor APC is an attenuator of spindle-pulling forces during C. elegans asymmetric cell division.Proc Natl Acad Sci U S A. 2018 Jan 30;115(5):E954-E963. doi: 10.1073/pnas.1712052115. Epub 2018 Jan 18. Proc Natl Acad Sci U S A. 2018. PMID: 29348204 Free PMC article.
-
Analysis of Haemonchus embryos at single cell resolution identifies two eukaryotic elongation factors as intervention target candidates.Comput Struct Biotechnol J. 2024 Jan 17;23:1026-1035. doi: 10.1016/j.csbj.2024.01.008. eCollection 2024 Dec. Comput Struct Biotechnol J. 2024. PMID: 38435301 Free PMC article.
-
A cytokinetic ring-driven cell rotation achieves Hertwig's rule in early development.Proc Natl Acad Sci U S A. 2024 Jun 18;121(25):e2318838121. doi: 10.1073/pnas.2318838121. Epub 2024 Jun 13. Proc Natl Acad Sci U S A. 2024. PMID: 38870057 Free PMC article.
-
Two populations of cytoplasmic dynein contribute to spindle positioning in C. elegans embryos.J Cell Biol. 2017 Sep 4;216(9):2777-2793. doi: 10.1083/jcb.201607038. Epub 2017 Jul 24. J Cell Biol. 2017. PMID: 28739679 Free PMC article.
References
-
- Afshar K, Willard FS, Colombo K, Johnston CA, McCudden CR, Siderovski DP, Gonczy P. RIC-8 is required for GPR-1/2-dependent Gα function during asymmetric division of C. elegans embryos. Cell. 2004;119:219–230. - PubMed
-
- Afshar K, Willard FS, Colombo K, Siderovski DP, Gonczy P. Cortical localization of the Gα protein GPA-16 requires RIC-8 function during C. elegans asymmetric cell division. Development. 2005;132:4449–4459. - PubMed
-
- Bergmann DC, Lee M, Robertson B, Tsou MF, Rose LS, Wood WB. Embryonic handedness choice in C. elegans involves the Gα protein GPA-16. Development. 2003;130:5731–5740. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
