

Extensive polymerase pausing during Drosophila axis patterning enables high-level and pliable transcription
- PMID: 23699410
- PMCID: PMC3672648
- DOI: 10.1101/gad.215459.113
Extensive polymerase pausing during Drosophila axis patterning enables high-level and pliable transcription
Abstract
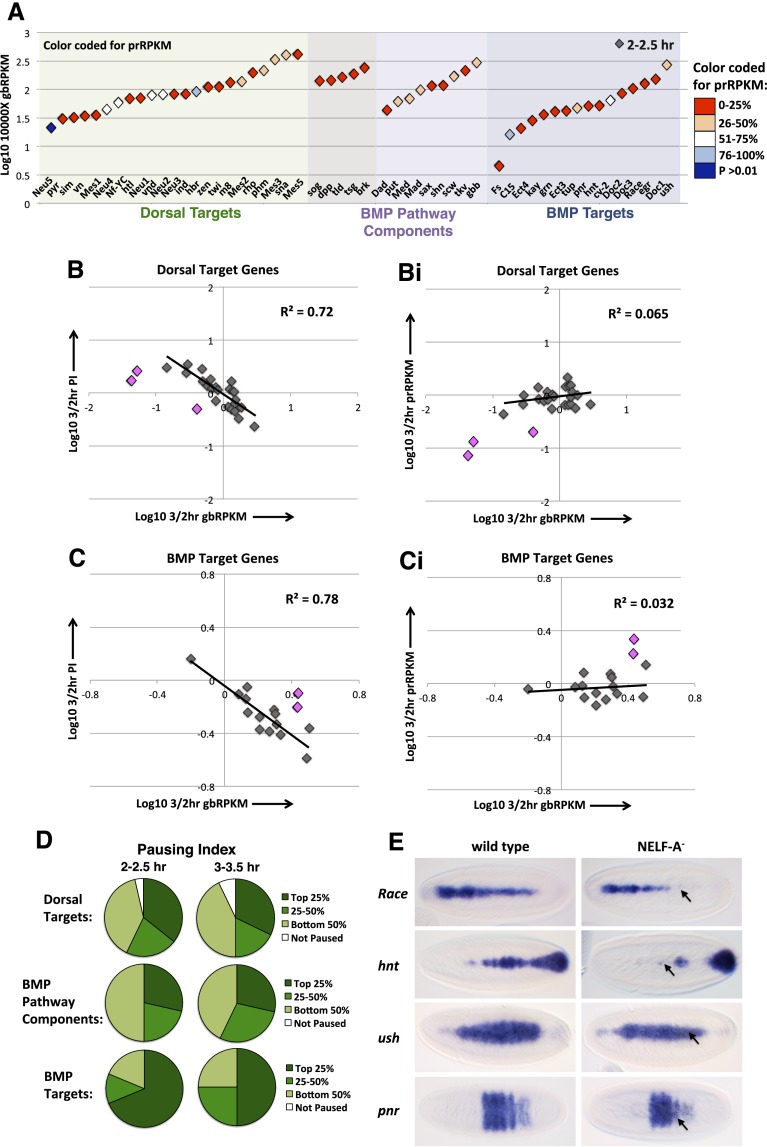
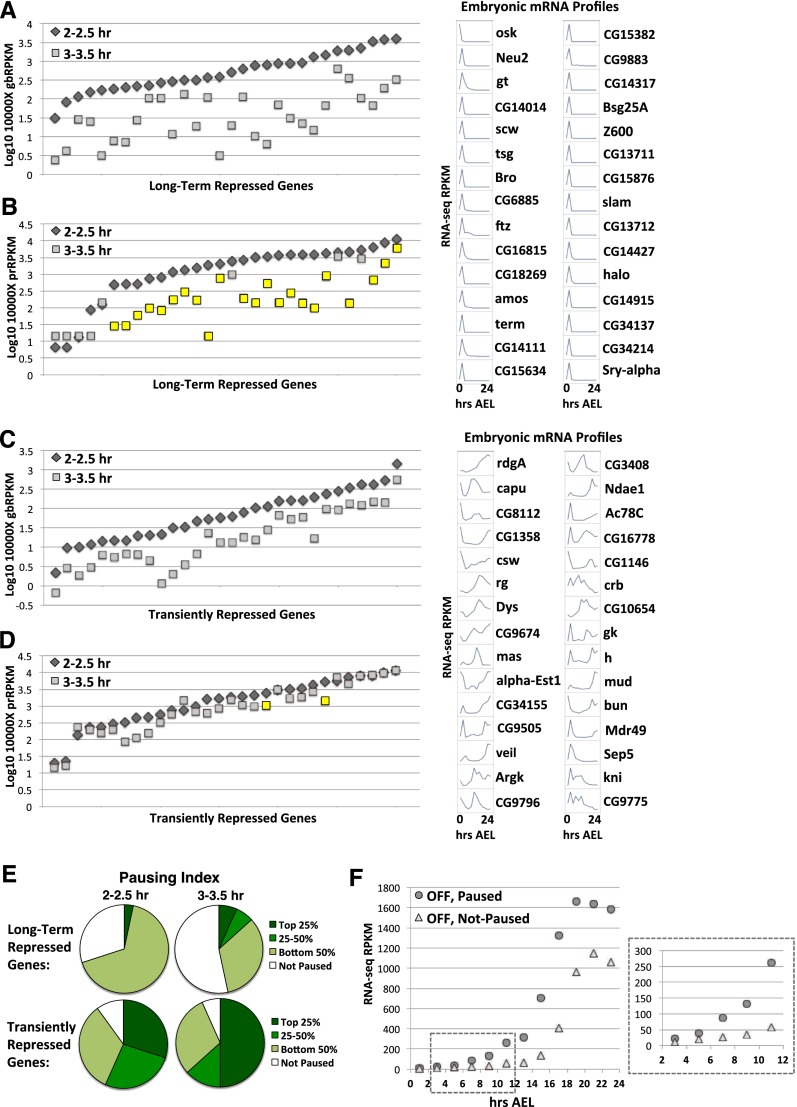
Cascades of zygotic gene expression pattern the anterior-posterior (AP) and dorsal-ventral (DV) axes of the early Drosophila embryo. Here, we used the global run-on sequencing assay (GRO-seq) to map the genome-wide RNA polymerase distribution during early Drosophila embryogenesis, thus providing insights into how genes are regulated. We identify widespread promoter-proximal pausing yet show that the presence of paused polymerase does not necessarily equate to direct regulation through pause release to productive elongation. Our data reveal that a subset of early Zelda-activated genes is regulated at the level of polymerase recruitment, whereas other Zelda target and axis patterning genes are predominantly regulated through pause release. In contrast to other signaling pathways, we found that bone morphogenetic protein (BMP) target genes are collectively more highly paused than BMP pathway components and show that BMP target gene expression requires the pause-inducing negative elongation factor (NELF) complex. Our data also suggest that polymerase pausing allows plasticity in gene activation throughout embryogenesis, as transiently repressed and transcriptionally silenced genes maintain and lose promoter polymerases, respectively. Finally, we provide evidence that the major effect of pausing is on the levels, rather than timing, of transcription. These data are discussed in terms of the efficiency of transcriptional activation required across cell populations during developmental time constraints.
Keywords: BMP; Dpp signaling pathway; Drosophila embryo; GRO-seq; axis patterning; promoter-proximal pausing; zygotic transcription.
Figures



Comment in
-
Gene expression: Time flies thanks to Pol II pausing.Nat Rev Genet. 2013 Jul;14(7):441. doi: 10.1038/nrg3514. Nat Rev Genet. 2013. PMID: 23752796 No abstract available.
References
-
- Arbeitman MN, Furlong EE, Imam F, Johnson E, Null BH, Baker BS, Krasnow MA, Scott MP, Davis RW, White KP 2002. Gene expression during the life cycle of Drosophila melanogaster. Science 297: 2270–2275 - PubMed
-
- Arora K, Levine MS, O'Connor MB 1994. The screw gene encodes a ubiquitously expressed member of the TGF-β family required for specification of dorsal cell fates in the Drosophila embryo. Genes Dev 8: 2588–2601 - PubMed
-
- Bentley DL, Groudine M 1986. A block to elongation is largely responsible for decreased transcription of c-myc in differentiated HL60 cells. Nature 321: 702–706 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases