

Advances in diffusion MRI acquisition and processing in the Human Connectome Project
- PMID: 23702418
- PMCID: PMC3720790
- DOI: 10.1016/j.neuroimage.2013.05.057
Advances in diffusion MRI acquisition and processing in the Human Connectome Project
Abstract
The Human Connectome Project (HCP) is a collaborative 5-year effort to map human brain connections and their variability in healthy adults. A consortium of HCP investigators will study a population of 1200 healthy adults using multiple imaging modalities, along with extensive behavioral and genetic data. In this overview, we focus on diffusion MRI (dMRI) and the structural connectivity aspect of the project. We present recent advances in acquisition and processing that allow us to obtain very high-quality in-vivo MRI data, whilst enabling scanning of a very large number of subjects. These advances result from 2 years of intensive efforts in optimising many aspects of data acquisition and processing during the piloting phase of the project. The data quality and methods described here are representative of the datasets and processing pipelines that will be made freely available to the community at quarterly intervals, beginning in 2013.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures

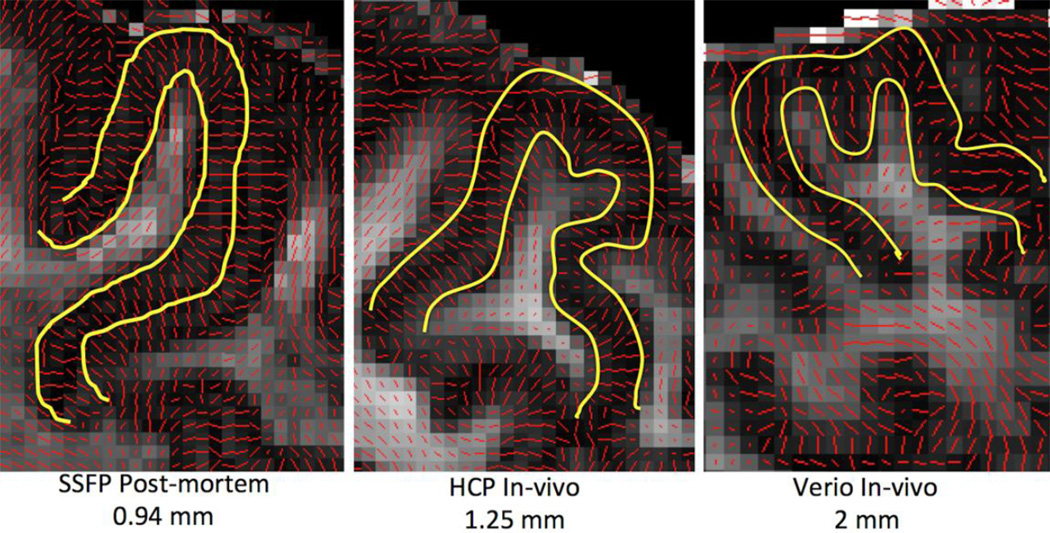
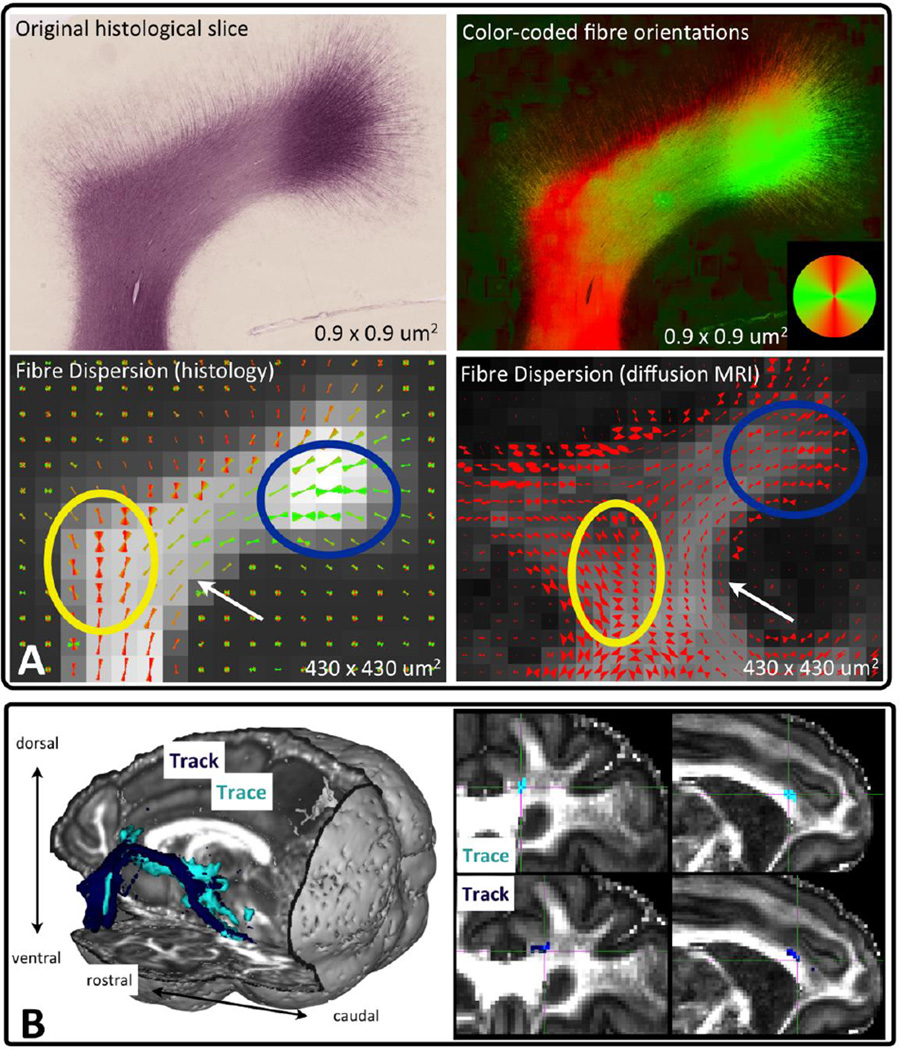
Similar articles
-
The WU-Minn Human Connectome Project: an overview.Neuroimage. 2013 Oct 15;80:62-79. doi: 10.1016/j.neuroimage.2013.05.041. Epub 2013 May 16. Neuroimage. 2013. PMID: 23684880 Free PMC article. Review.
-
The minimal preprocessing pipelines for the Human Connectome Project.Neuroimage. 2013 Oct 15;80:105-24. doi: 10.1016/j.neuroimage.2013.04.127. Epub 2013 May 11. Neuroimage. 2013. PMID: 23668970 Free PMC article.
-
Adding dynamics to the Human Connectome Project with MEG.Neuroimage. 2013 Oct 15;80:190-201. doi: 10.1016/j.neuroimage.2013.05.056. Epub 2013 May 20. Neuroimage. 2013. PMID: 23702419 Free PMC article.
-
The Lifespan Human Connectome Project in Aging: An overview.Neuroimage. 2019 Jan 15;185:335-348. doi: 10.1016/j.neuroimage.2018.10.009. Epub 2018 Oct 15. Neuroimage. 2019. PMID: 30332613 Free PMC article.
-
The Human Connectome Project: a data acquisition perspective.Neuroimage. 2012 Oct 1;62(4):2222-31. doi: 10.1016/j.neuroimage.2012.02.018. Epub 2012 Feb 17. Neuroimage. 2012. PMID: 22366334 Free PMC article. Review.
Cited by
-
Gradual changes in microarchitectural properties of cortex and juxtacortical white matter: Observed by anatomical and diffusion MRI.Magn Reson Med. 2022 Dec;88(6):2485-2503. doi: 10.1002/mrm.29413. Epub 2022 Aug 31. Magn Reson Med. 2022. PMID: 36045582 Free PMC article.
-
Group-wise consistent cortical parcellation based on connectional profiles.Med Image Anal. 2016 Aug;32:32-45. doi: 10.1016/j.media.2016.02.009. Epub 2016 Mar 14. Med Image Anal. 2016. PMID: 27054276 Free PMC article.
-
Diffusion Imaging in the Post HCP Era.J Magn Reson Imaging. 2021 Jul;54(1):36-57. doi: 10.1002/jmri.27247. Epub 2020 Jun 20. J Magn Reson Imaging. 2021. PMID: 32562456 Free PMC article. Review.
-
Automated segmentation of the hypothalamus and associated subunits in brain MRI.Neuroimage. 2020 Dec;223:117287. doi: 10.1016/j.neuroimage.2020.117287. Epub 2020 Aug 25. Neuroimage. 2020. PMID: 32853816 Free PMC article.
-
Assessment of the Depiction of Superficial White Matter Using Ultra-High-Resolution Diffusion MRI.Hum Brain Mapp. 2024 Oct;45(14):e70041. doi: 10.1002/hbm.70041. Hum Brain Mapp. 2024. PMID: 39392220 Free PMC article.
References
-
- Alexander DC. Maximum entropy spherical deconvolution for diffusion MRI. Inf Process Med Imaging. 2005;19:76–87. - PubMed
-
- Alexander DC, Hubbard PL, Hall MG, Moore EA, Ptito M, Parker GJ, Dyrby TB. Orientationally invariant indices of axon diameter and density from diffusion MRI. Neuroimage. 2010;52:1374–1389. - PubMed
-
- Anderson AW. Measurement of fiber orientation distributions using high angular resolution diffusion imaging. Magn Reson Med. 2005;54:1194–1206. - PubMed
-
- Andersson JLR, Xu J, Yacoub E, Auerbach EJ, Moeller S, Ugurbil K. A comprehensive gaussian process framework for correcting distortions and movements in diffusion images; Melbourne, Australia. International Society for Magnetic Resonance in Medicine (ISMRM) Annual Meeting; 2012. p. 2426.
Publication types
MeSH terms
Grants and funding
- 1U54MH091657-01/MH/NIMH NIH HHS/United States
- 098369/WT_/Wellcome Trust/United Kingdom
- R01 MH060974/MH/NIMH NIH HHS/United States
- P30 NS076408/NS/NINDS NIH HHS/United States
- S10 RR022984/RR/NCRR NIH HHS/United States
- 1S10RR022984-01A1/RR/NCRR NIH HHS/United States
- R01 MH60974/MH/NIMH NIH HHS/United States
- P41 EB015894/EB/NIBIB NIH HHS/United States
- F30 MH097312/MH/NIMH NIH HHS/United States
- U54 MH091657/MH/NIMH NIH HHS/United States
- P30 NS057091/NS/NINDS NIH HHS/United States
- 090955/WT_/Wellcome Trust/United Kingdom
- G0800578/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
