

Review
doi: 10.1038/cdd.2013.50.
Epub 2013 May 24.
Glycobiology of cell death: when glycans and lectins govern cell fate
Affiliations
- PMID: 23703323
- PMCID: PMC3705604
- DOI: 10.1038/cdd.2013.50
Item in Clipboard
Review
Glycobiology of cell death: when glycans and lectins govern cell fate
Cell Death Differ.
2013 Aug.
Abstract
Although one typically thinks of carbohydrates as associated with cell growth and viability, glycosylation also has an integral role in many processes leading to cell death. Glycans, either alone or complexed with glycan-binding proteins, can deliver intracellular signals or control extracellular processes that promote initiation, execution and resolution of cell death programs. Herein, we review the role of glycans and glycan-binding proteins as essential components of the cell death machinery during physiologic and pathologic settings.
Figures
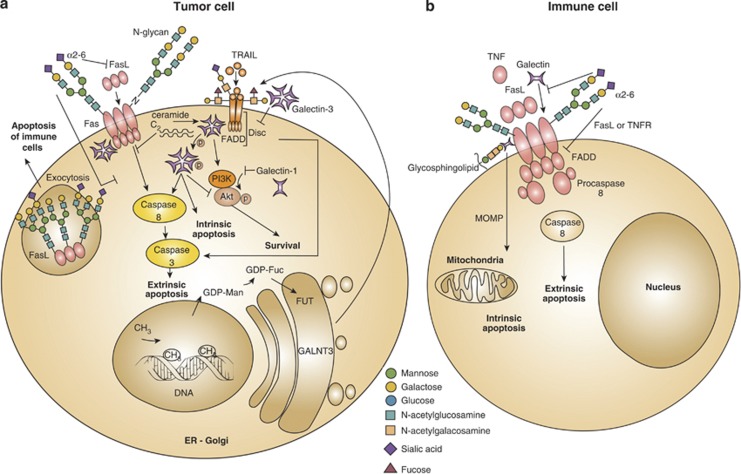
Influence of glycans and glycan-binding proteins in death receptor signaling and function. In tumor cells, alterations of the components comprising the ‘glycosylation machinery' generate aberrant O-glycans and N-glycans on Fas and TRAIL receptors, which modulate apoptosis. Upregulation of GALNT3, FUT and GDP-FUC as a result of DNA methylation increases tumor cell sensitivity to extrinsic apoptosis through TRAIL. In addition, α2-6-linked sialic acid decorating Fas receptor on tumor cells blocks Fas ligand internalization, Fas–FADD complex formation and activation of caspase-8 and -3, thus attenuating cell death via the extrinsic pathway. Intracellular and extracellular galectins can modulate survival and apoptotic signaling pathways in tumor cells. Intracellular galectin-1 decreases Akt activity and induces apoptosis. By contrast, intracellular galectin-3 either free or associated with C2-ceramide cooperates with PI3K/Akt to increase tumor cell survival. However, in some cases, intracellular galectin-3 promotes extrinsic and intrinsic apoptosis, either through association with Fas or via phosphorylation at serine 6. Extracellular galectin-3 inhibits the extrinsic apoptosis cascade by anchoring TRAIL receptors through glycosylation-dependent mechanisms. Moreover, glycosylation of intracellular Fas ligands in tumor cells facilitates ligand secretion as a strategy of tumor-immune escape (a). Fas receptor on immune cell bears N-glycans to stabilize complexes like DISC, generated during the execution of the extrinsic apoptotic pathway. Increased α2-6 sialylation of N-glycans impairs Fas internalization, FADD–Fas complex formation, activation of caspase-8 and -3 and T-cell homeostasis. α2-6 sialylation further interferes with galectin-1 binding to glycoproteins (e.g. CD45, CD43), and also interrupts ligand binding to TNFR1 on macrophages and decreases apoptosis. Galectin-1 interacts with Fas on resting T cells and stimulates both the intrinsic and extrinsic apoptotic signaling routes (b). GALNT3, N-acetyl galactosyltransferase-3; FUT, fucosyltransferase; GDP-FUC, guanosine diphosphate-fucose; Akt, serine/threonine-specific protein kinase; Ser6, serine at position 6; CD45, protein tyrosine phosphatase; CD43; sialophorin
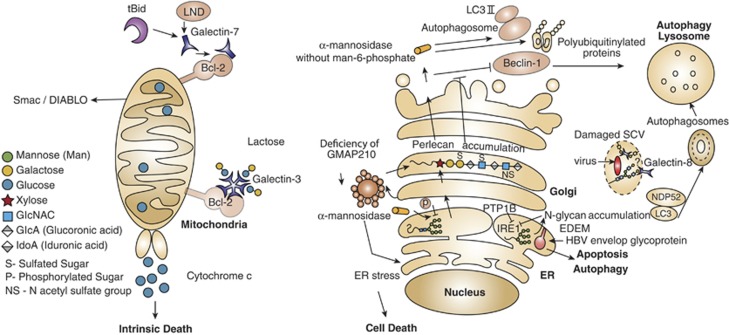
Glycans and glycan-binding proteins are integral components of the autophagy and apoptosis machineries. Interaction of galectins with various intracellular proteins either in a glycan-dependent or -independent manner may control cell death in diverse subcellular compartments. In tBid- or LND-treated mitochondria, as well as in untreated mitochondria, intracellular galectin-3 and galectin-7 form heterodimers with Bcl-2, which modulate cellular apoptosis. Galectin-7–Bcl-2 complex promotes release of cytochrome c and Smac/DIABLO factors, facilitating the intrinsic death pathway. In contrast, endogenous galectin-8 is a cytoprotective intermediate protein that restrains Salmonella proliferation and inflammation by neutralizing bacteria-damaged SCV in human host cells. Host galectin-8 binds glycans expressed on the SCV, recruits and forms complexes with NDP52 and LC3, which lead to encapsulating SCV into an autophagosome. Finally, the unwanted cargo is degraded in the lysosome. It is not clear whether galectin-8 binds to N-glycans or to O-glycans of the SCV. The cargo proteins, mannosidase-like protein (EDEM) in the ER and hydrolase α-mannosidase, as well as accumulation of N-glycans in the ER are involved in cell death. Whereas EDEM facilitates the removal of envelope glycoprotein of hepatitis B virus (HBV)-infected cells in an autophagy-dependent manner, the absence of the phosphatase PTP-1B impairs EDEM transcription and suppresses apoptosis. Defective mannose-6 phosphate targeting mechanism impairs transportation of α-mannosidase and of many glycan-metabolizing enzymes from the secretory pathway to the lysosome. Such impairment causes the accumulation of polyubiquitinylated protein aggregates and LC3-II and unchanged expression of Beclin-1, which together inhibit autophagy. Golgi architecture protein GMAP210 has been also involved in cell death signaling programs. Deficiency of this protein promotes ER stress accompanied by abnormal glycosylation, accumulation of perlican in the Golgi and cell death. Whether intermediates of Golgi glycosylation are involved in cell death is still not clear. tBid, truncated Bcl-2 homology-3 (BH3)-interacting domain death agonist; LND, lonidamine; Smac/DIABLO, second-mitochondria-derived activator of caspases; SCV, Salmonella-containing vacuole; EDEM, ER degradation-enhancing α-mannosidase-like protein; PTP-1B, protein-tyrosine phosphatase 1B; GMAP210, Golgi-associated microtubule-binding protein 210
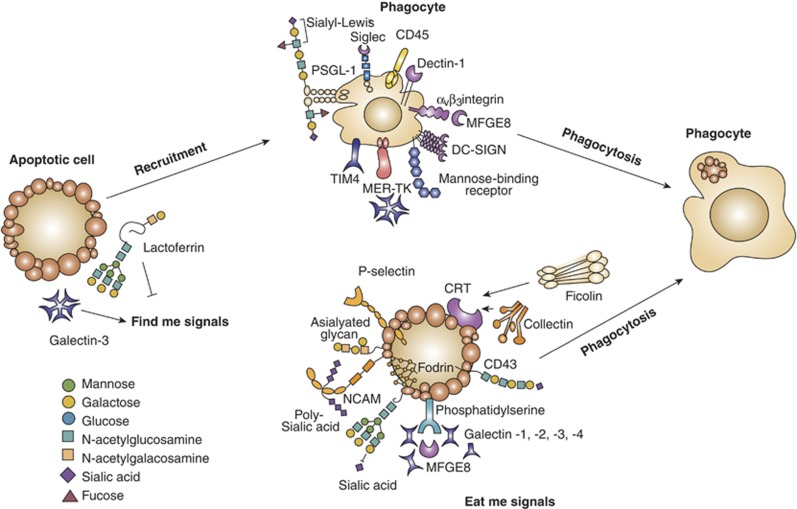
Glycans and lectins in the resolving stages of cell death. Removal of apoptotic cells involves ‘find me' signals, ‘eat me' signals and engulfment of dying cells by phagocytes. Selected lectin–glycan recognition systems, which have been identified as part of that apoptotic cell removal, are shown. Among the ‘find me' and ‘eat me' signals illustrated, galectins and soluble glycoproteins are involved in the initiation of phagocytosis and tethering. Galactose-terminated glycans and also poly-sialic acids act as ‘eat me' signals, whereas sialy-Lewisx terminal groups have been involved in tethering. P-selectin and calreticulin displayed on apoptotic cells have essential roles in the initiation of phagocytosis and C-type lectins and siglecs have been implicated in tethering. TIM-4, T-cell immunoglobulin mucin-4; PSGL-1, P-selectin glycoprotein ligand-1; DC-SIGN, dendritic cell-specific intracellular adhesion molecule-3-garbbing non-integrin; NCAM, neural cell adhesion molecule
Similar articles
-
Glycobiology of cellular expiry: Decrypting the role of glycan-lectin regulatory complex and therapeutic strategies focusing on cancer.Biochem Pharmacol. 2023 Jan;207:115367. doi: 10.1016/j.bcp.2022.115367. Epub 2022 Dec 5. Biochem Pharmacol. 2023. PMID: 36481348 Review.
-
Glycobiology of the ocular surface: mucins and lectins.Jpn J Ophthalmol. 2013 Mar;57(2):150-5. doi: 10.1007/s10384-012-0228-2. Epub 2013 Jan 17. Jpn J Ophthalmol. 2013. PMID: 23325272 Free PMC article. Review.
-
"Stuck on sugars - how carbohydrates regulate cell adhesion, recognition, and signaling".Glycoconj J. 2019 Aug;36(4):241-257. doi: 10.1007/s10719-019-09876-0. Epub 2019 Jul 2. Glycoconj J. 2019. PMID: 31267247 Free PMC article.
-
Glycobiology of immune responses.Ann N Y Acad Sci. 2012 Apr;1253:1-15. doi: 10.1111/j.1749-6632.2012.06492.x. Ann N Y Acad Sci. 2012. PMID: 22524422 Free PMC article. Review.
-
Glycobiology in the cytosol: the bitter side of a sweet world.Biochim Biophys Acta. 2009 Feb;1790(2):81-94. doi: 10.1016/j.bbagen.2008.09.009. Epub 2008 Oct 8. Biochim Biophys Acta. 2009. PMID: 18952151 Review.
Cited by
-
Harnessing benefit from targeting tumor associated carbohydrate antigens.Hum Vaccin Immunother. 2017 Feb;13(2):323-331. doi: 10.1080/21645515.2017.1264789. Hum Vaccin Immunother. 2017. PMID: 27929800 Free PMC article. Review.
-
Hallmarks of glycosylation in cancer.Oncotarget. 2016 Jun 7;7(23):35478-89. doi: 10.18632/oncotarget.8155. Oncotarget. 2016. PMID: 27007155 Free PMC article. Review.
-
Multi-targeted therapy resistance via drug-induced secretome fucosylation.Elife. 2023 Mar 24;12:e75191. doi: 10.7554/eLife.75191. Elife. 2023. PMID: 36961502 Free PMC article.
-
Galectin-9 in Gastroenterological Cancer.Int J Mol Sci. 2023 Mar 24;24(7):6174. doi: 10.3390/ijms24076174. Int J Mol Sci. 2023. PMID: 37047155 Free PMC article. Review.
-
Molecular mechanisms of cell death: recommendations of the Nomenclature Committee on Cell Death 2018.Cell Death Differ. 2018 Mar;25(3):486-541. doi: 10.1038/s41418-017-0012-4. Epub 2018 Jan 23. Cell Death Differ. 2018. PMID: 29362479 Free PMC article. Review.
References
-
- Lockshin RA, Williams CM. Programmed cell death-I. Cytology of degeneration in the intersegmental muscles of the pernyi silkmoth. J Insect Physiol. 1965;11:123–133. - PubMed
-
- Sharon N, Lis H. History of lectins: from hemagglutinins to biological recognition molecules. Glycobiology. 2004;14:53R–62R. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
