

Primuline derivatives that mimic RNA to stimulate hepatitis C virus NS3 helicase-catalyzed ATP hydrolysis
- PMID: 23703611
- PMCID: PMC3707695
- DOI: 10.1074/jbc.M113.463166
Primuline derivatives that mimic RNA to stimulate hepatitis C virus NS3 helicase-catalyzed ATP hydrolysis
Abstract
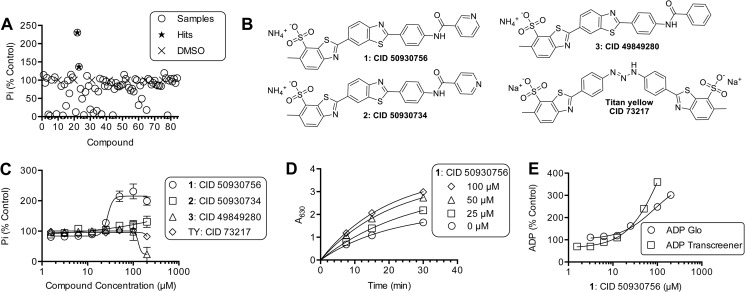
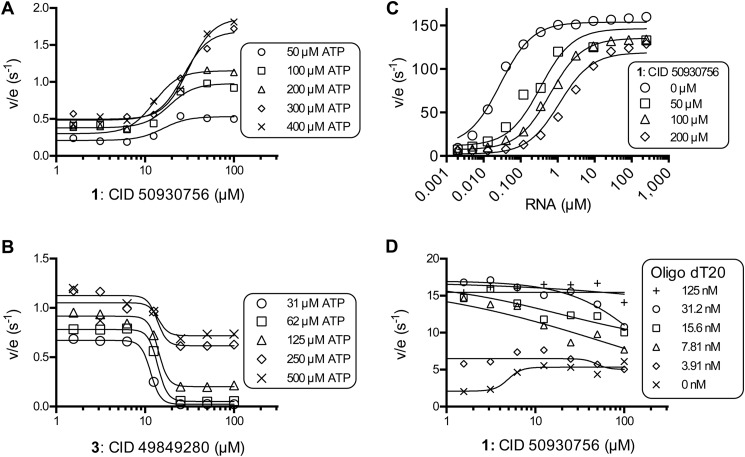
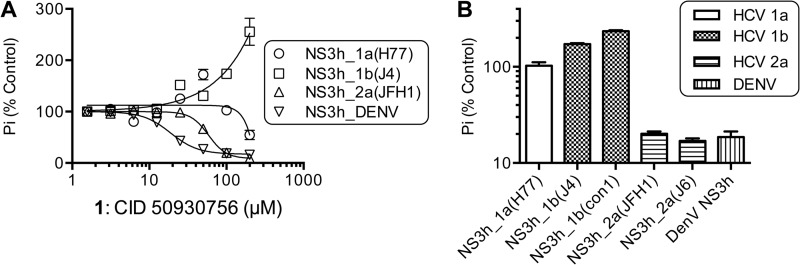
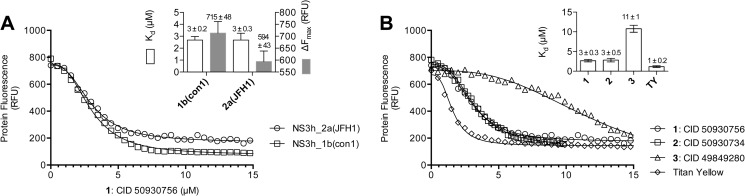
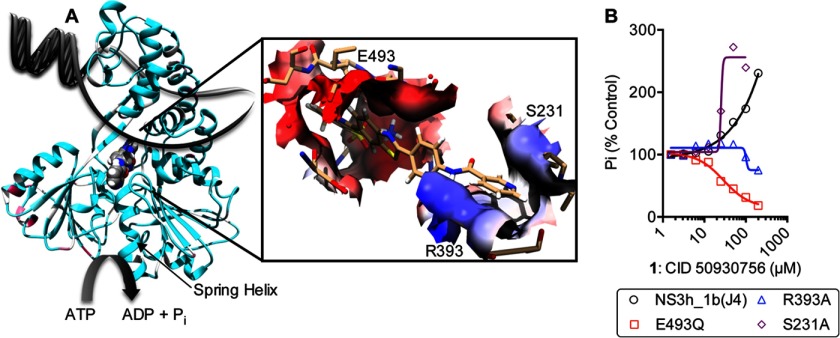
ATP hydrolysis fuels the ability of helicases and related proteins to translocate on nucleic acids and separate base pairs. As a consequence, nucleic acid binding stimulates the rate at which a helicase catalyzes ATP hydrolysis. In this study, we searched a library of small molecule helicase inhibitors for compounds that stimulate ATP hydrolysis catalyzed by the hepatitis C virus (HCV) NS3 helicase, which is an important antiviral drug target. Two compounds were found that stimulate HCV helicase-catalyzed ATP hydrolysis, both of which are amide derivatives synthesized from the main component of the yellow dye primuline. Both compounds possess a terminal pyridine moiety, which was critical for stimulation. Analogs lacking a terminal pyridine inhibited HCV helicase catalyzed ATP hydrolysis. Unlike other HCV helicase inhibitors, the stimulatory compounds differentiate between helicases isolated from various HCV genotypes and related viruses. The compounds only stimulated ATP hydrolysis catalyzed by NS3 purified from HCV genotype 1b. They inhibited helicases from other HCV genotypes (e.g. 1a and 2a) or related flaviviruses (e.g. Dengue virus). The stimulatory compounds interacted with HCV helicase in the absence of ATP with dissociation constants of about 2 μM. Molecular modeling and site-directed mutagenesis studies suggest that the stimulatory compounds bind in the HCV helicase RNA-binding cleft near key residues Arg-393, Glu-493, and Ser-231.
Keywords: ATPases; Chemical Biology; Drug Discovery; High Throughput Screening (HTS); Molecular Motors; RNA Helicase.
Figures
Similar articles
-
Fluorescent primuline derivatives inhibit hepatitis C virus NS3-catalyzed RNA unwinding, peptide hydrolysis and viral replicase formation.Antiviral Res. 2012 Nov;96(2):245-55. doi: 10.1016/j.antiviral.2012.08.006. Epub 2012 Aug 25. Antiviral Res. 2012. PMID: 22940425 Free PMC article.
-
Hepatitis C virus NS3 ATPases/helicases from different genotypes exhibit variations in enzymatic properties.J Virol. 2003 Apr;77(7):3950-61. doi: 10.1128/jvi.77.7.3950-3961.2003. J Virol. 2003. PMID: 12634355 Free PMC article.
-
Mechanism and specificity of a symmetrical benzimidazolephenylcarboxamide helicase inhibitor.Biochemistry. 2010 Mar 9;49(9):1822-32. doi: 10.1021/bi901974a. Biochemistry. 2010. PMID: 20108979 Free PMC article.
-
Structure and function of hepatitis C virus NS3 helicase.Curr Top Microbiol Immunol. 2000;242:171-96. doi: 10.1007/978-3-642-59605-6_9. Curr Top Microbiol Immunol. 2000. PMID: 10592661 Review.
-
Step-by-step progress toward understanding the hepatitis C virus RNA helicase.Hepatology. 2006 Jun;43(6):1392-5. doi: 10.1002/hep.21200. Hepatology. 2006. PMID: 16729317 Free PMC article. Review.
Cited by
-
Benzothiazole and Pyrrolone Flavivirus Inhibitors Targeting the Viral Helicase.ACS Infect Dis. 2015 Mar 13;1(3):140-148. doi: 10.1021/id5000458. ACS Infect Dis. 2015. PMID: 26029739 Free PMC article.
-
Simultaneously Targeting the NS3 Protease and Helicase Activities for More Effective Hepatitis C Virus Therapy.ACS Chem Biol. 2015 Aug 21;10(8):1887-96. doi: 10.1021/acschembio.5b00101. Epub 2015 May 22. ACS Chem Biol. 2015. PMID: 25961497 Free PMC article.
-
Flavivirus enzymes and their inhibitors.Enzymes. 2021;49:265-303. doi: 10.1016/bs.enz.2021.07.006. Epub 2021 Sep 1. Enzymes. 2021. PMID: 34696835 Free PMC article.
-
Role of the Conserved DECH-Box Cysteine in Coupling Hepatitis C Virus Helicase-Catalyzed ATP Hydrolysis to RNA Unwinding.Biochemistry. 2018 Oct 30;57(43):6247-6255. doi: 10.1021/acs.biochem.8b00796. Epub 2018 Oct 16. Biochemistry. 2018. PMID: 30281972 Free PMC article.
-
Motif V regulates energy transduction between the flavivirus NS3 ATPase and RNA-binding cleft.J Biol Chem. 2020 Feb 7;295(6):1551-1564. doi: 10.1074/jbc.RA119.011922. Epub 2019 Dec 30. J Biol Chem. 2020. PMID: 31914411 Free PMC article.
References
-
- Li K., Frankowski K. J., Belon C. A., Neuenswander B., Ndjomou J., Hanson A. M., Shanahan M. A., Schoenen F. J., Blagg B. S., Aubé J., Frick D. N. (2012) Optimization of potent hepatitis C virus NS3 helicase inhibitors isolated from the yellow dyes thioflavine S and primuline. J. Med. Chem. 55, 3319–3330 - PMC - PubMed
-
- Bacon B. R., Gordon S. C., Lawitz E., Marcellin P., Vierling J. M., Zeuzem S., Poordad F., Goodman Z. D., Sings H. L., Boparai N., Burroughs M., Brass C. A., Albrecht J. K., Esteban R. (2011) Boceprevir for previously treated chronic HCV genotype 1 infection. N. Engl. J. Med. 364, 1207–1217 - PMC - PubMed
-
- Zeuzem S., Andreone P., Pol S., Lawitz E., Diago M., Roberts S., Focaccia R., Younossi Z., Foster G. R., Horban A., Ferenci P., Nevens F., Müllhaupt B., Pockros P., Terg R., Shouval D., van Hoek B., Weiland O., Van Heeswijk R., De Meyer S., Luo D., Boogaerts G., Polo R., Picchio G., Beumont M. (2011) Telaprevir for retreatment of HCV infection. N. Engl. J. Med. 364, 2417–2428 - PubMed
-
- Kim J. L., Morgenstern K. A., Griffith J. P., Dwyer M. D., Thomson J. A., Murcko M. A., Lin C., Caron P. R. (1998) Hepatitis C virus NS3 RNA helicase domain with a bound oligonucleotide: the crystal structure provides insights into the mode of unwinding. Structure 6, 89–100 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
